Co2+对红霉素生物合成过程的影响
2007-03-31 20:56:40 来源:华东理工大学学报(自然科学版) 评论:0 点击:
随着分子生物学和生物技术的发展,人们对红霉素生物合成过程和生物合成基因的认识已经比较清楚,但红霉素生物合成过程中所涉及到的酶对发酵过程作用的相关报道较少。红霉素属次级代谢产物,而菌体的初级代谢对次级代谢有着极大的影响。因此测定糖代谢过程中的一些关键酶能更好地了解红霉素产生菌的代谢过程
红霉素属14元环大环内酯类抗生素,其内酯环是由1分子丙酰CoA和6分子甲基丙二酰CoA经缩合、酮还原、脱水和烯还原等多轮循环完成。在大环内酯抗生素生物合成中,内酯环的形成往往是限制性步骤,因此丙酰CoA和甲基丙二酰CoA的合成显得极为重要。而在红霉素合成中,丙酰CoA也可由甲基丙二酰CoA脱羧得到,因而甲基丙二酰CoA的合成是关键。目前认为,红霉素合成中的甲基丙二酰CoA主要由琥珀酰CoA通过甲基丙二酰CoA异构酶转化而来[1]。甲基丙二酰CoA异构酶需要5′-腺苷钴胺素作为辅因子,而钴离子又是其中心金属离子,因此在培养基中加入Co2+可能提高该酶的活性,从而对红霉素合成有促进作用。本文考察了Co2+对发酵的影响,同时测定了甲基丙二酰CoA异构酶以及糖代谢过程中酶的时序变化,以及三羧酸循环中部分有机酸的含量变化,对红霉素发酵在细胞水平上有了一个新的认识。
1.1 菌种和培养基菌种:
红霉素链霉菌,由本课题组自备。斜面培养基(g/L):可溶性淀粉10,硫酸铵3,氯化钠3,玉米浆13,碳酸钙2.5,琼脂粉10,pH=7.0~7.2。种子培养基(g/L):淀粉20,糊精25,玉米浆30,黄豆饼粉25,硫酸铵1.8,氯化钠4,硝酸铵1.2,碳酸钙2,豆油2,pH=7.0~7.2。摇瓶培养基(g/L):淀粉50,糊精40,玉米浆4,黄豆饼粉30,硫酸铵1.5,氯化钠2,碳酸钙4,豆油2,pH=7.0~7.2。发酵罐培养基(g/L):淀粉39,糊精6,玉米浆27,黄豆饼粉22,硫酸铵1.27,氯化钠1.7,碳酸钙3.3,豆油6.6,pH=7.0~7.2。
1.2 发酵培养方法
1.2.1 摇瓶发酵 种子培养:将斜面孢子挖块1cm×1cm接种在种子培养基中,在摇床34°C培养44~48h,摇床转速为220~240r/min。摇瓶培养:将种子液(接种量为10%)接入摇瓶培养基中,在摇床34°C培养约160h,摇床转速同上。
1.2.2 发酵罐发酵 一级种子培养:同摇瓶发酵中的种子培养。二级种子培养:将一级种子液(接种量为5%)接入种子培养基中,在摇床34°C培养24h,摇床转速同上。发酵罐培养:将二级种子液(接种量为8%)接入发酵罐培养基中,34°C培养8h后降温至33°C。控制发酵液溶氧在30%以上,总糖含量在30~33g/L,pH在7.0左右。
1.3 发酵过程参数检测还原糖和总糖的测定:取不同时期的培养液,用费林法测定其中的还原糖和总糖。
氨基氮的测定:取不同时期的培养液,用甲醛法测定其中的氨基氮含量。红霉素化学效价测定:采用磷酸法。即取发酵滤液稀释到200~300U/mL,吸取10mol/L磷酸4mL于10mL的容量瓶,加入0.8mL的稀释液,沸水浴3min。冷却后,用10mol/L磷酸定容至刻度线,在485nm处测吸光度。菌浓测定:采用离心压缩细胞体积法(PVC)。取10mL发酵液,2500r/min离心10min,测量细胞等固形物所占总体积的分数(φCell)即表示为菌浓。蛋白质浓度测定:考马斯亮蓝法[2],以牛血清蛋白作为测定参照标准。己糖激酶(HK)比活力的测定:见文献[3]。6-磷酸葡萄糖脱氢酶(G6PDH)比活力的测定:见文献[4]。磷酸果糖激酶(PFK)比活力的测定:见文献[5]。异柠檬酸脱氢酶(ICD)比活力的测定:见文献[6]。甲基丙二酰CoA异构酶(MCM)比活力的测定:见文献[7]。甲基丙二酰CoA羧基转移酶(MCT)比活力的测定:见文献[8]。
2 实验结果
2.1 Co2+浓度的选择在摇瓶培养基中加入CoCl2,浓度分别为0、0.1、0.2、0.3g/L。发酵160h后放瓶,结果见表1。从表1可以看出,加入CoCl2对提高红霉素效价有明显的作用,其中添加0.2g/L的效果最好,比对照提高近30%。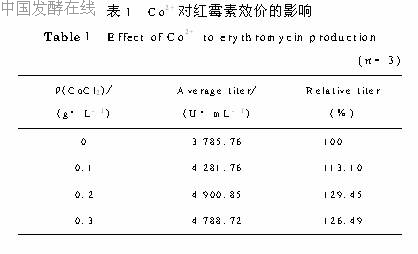
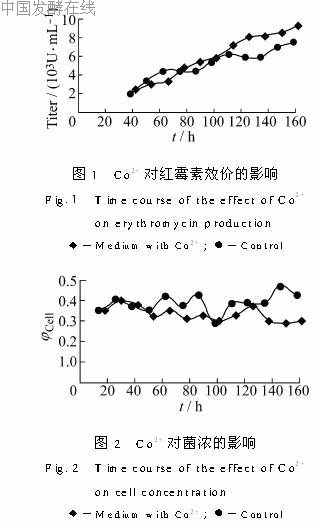
2.2 Co2+对红霉素效价的影响在摇瓶实验中,浓度为0.2g/L的Co2+提高红霉素效价最显著,故选定该浓度在发酵罐上做进一步实验。以不加CoCl2为对照。效价随时间的变化如图1所示。菌浓随时间的变化如图2所示。由图可以看出,加入Co2+使红霉素效价增长较快,在发酵100h后效价已明显高于对照。而菌浓在60h前基本上没有差别,在发酵中后期对照罐的菌浓比加Co2+的罐略高。
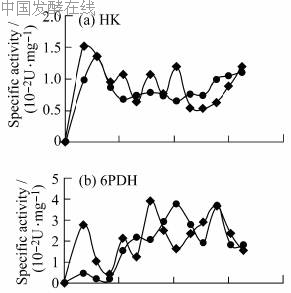
2.3 Co2+对糖代谢途径关键酶的影响为进一步了解Co2+在菌体代谢中可能起的作用,分别测定了糖代谢途径中糖激酶(HK)、6-磷酸葡萄糖脱氢酶(6PDH)、磷酸果糖激酶(PFK)和异柠檬酸脱氢酶(ICD)的比活,以不加CoCl2为对照,结果见图3。
HK是进入糖代谢的第一个限速酶,反映了葡萄糖进入细胞的通量。如图3a所示,在整个发酵过程中前期的比活较高,随后略有降低并维持在一定水平。这表明在生长阶段需要消耗大量葡萄糖来合成菌体,之后菌体转入次级代谢,需要的葡萄糖仅用于维持菌体生存,因而HK的比活有所下降。加入Co2+的比活比对照略高。
6PDH是HMP途径的限速酶,在发酵前50h比活都较低,50h后开始升高。18h时,加入Co2+的比活达到对照的6倍,可见Co2+的加入使得发酵前期HMP途径加强(图3b)。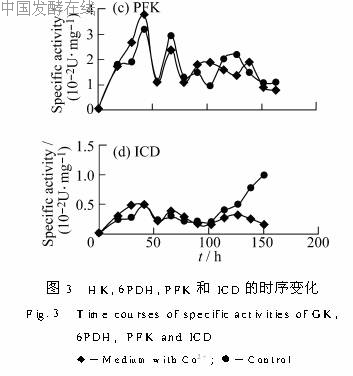
PFK是EMP途径中的关键酶,其比活趋势与HK较相似,在40h前较高,随后有所降低。加入Co2+的比活在前40h比对照略高,之后两者水平相差不大(图3c)。
ICD催化异柠檬酸生成α-酮戊二酸,此反应不可逆,是TCA循环中的限速步骤。如图3d所示,在30~40h和70h左右异柠檬酸脱氢酶各有一个比活高峰。加入Co2+,前期比活的变化趋势与对照相似,在125h后有下降趋势。
2.4 Co2+对甲基丙二酰CoA异构酶的影响甲基丙二酰CoA主要有3条合成途径:由丙酰CoA在丙酰CoA羧化酶的作用下生成;由丙酰CoA和丙酮酸通过甲基丙二酰CoA羧基转移酶(MCT)生成;由三羧酸循环的中间物琥珀酰CoA通过甲基丙二酰CoA异构酶(MCM)生成。Donadio等[9]曾报道,糖多孢链霉菌合成红霉素并不需要丙酰CoA羧化酶的活性。因此只测定了发酵过程中后2种酶(MCM、MCT)的比活,结果见图4。
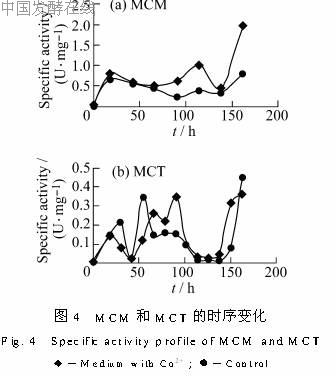
由图4a可看出,在对照中,甲基丙二酰CoA异构酶的比活在18h左右出现第一个高峰,随着发酵的进行比活有逐渐降低的趋势,90h后略有上升。加Co2+的比活始终比对照高。前期的变化趋势与对照相似,但在66h后比活就开始上升,在138h约为对照的2.5倍。这说明Co2+能显著的提高该酶比活。由图4b可看出,甲基丙二酰CoA羧基转移酶主要有2个比活高峰,在18h到30h菌体生长期比活逐渐升高,在42h迅速下降,54h又出现一个比活高峰。加Co2+的比活力第二个高峰与对照相比出现稍晚,但延续的时间较长,即在60~100h该酶的比活均处在较高的水平。可见Co2+对甲基丙二酰CoA羧基转移酶的合成有也一定促进作用。
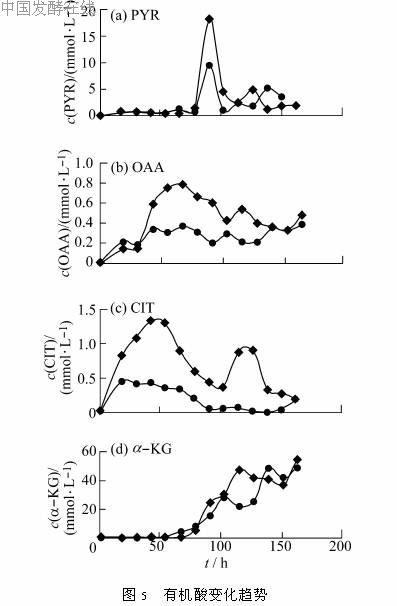
2.5 发酵过程中有机酸含量的变化趋势酶比活的变化反映的是细胞中存在的酶量,还不能充分体现各酶在细胞中受抑制或激活的实际情况,因此用HPLC测定了发酵液中的一些有机酸的含量,结合有机酸的变化可以对细胞的代谢途径有更深入的了解。有机酸的测定结果见图5。
由图4a可看出,在对照中,甲基丙二酰CoA异构酶的比活在18h左右出现第一个高峰,随着发酵的进行比活有逐渐降低的趋势,90h后略有上升。加Co2+的比活始终比对照高。前期的变化趋势与对照相似,但在66h后比活就开始上升,在138h约为对照的2.5倍。这说明Co2+能显著的提高该酶比活。由图4b可看出,甲基丙二酰CoA羧基转移酶主要有2个比活高峰,在18h到30h菌体生长期比活逐渐升高,在42h迅速下降,54h又出现一个比活高峰。加Co2+的比活力第二个高峰与对照相比出现稍晚,但延续的时间较长,即在60~100h该酶的比活均处在较高的水平。可见Co2+对甲基丙二酰CoA羧基转移酶的合成有也一定促进作用。2.5 发酵过程中有机酸含量的变化趋势酶比活的变化反映的是细胞中存在的酶量,还不能充分体现各酶在细胞中受抑制或激活的实际情况,因此用HPLC测定了发酵液中的一些有机酸的含量,结合有机酸的变化可以对细胞的代谢途径有更深入的了解。有机酸的测定结果见图5。
(α-KG)含量的变化趋势与柠檬酸相反,在78h前较低,之后有明显增加。加入Co2+后,α-酮戊二酸含量在90~130h比对照高,其余时间与对照基本相同。
3 讨 论
在摇瓶发酵培养基中加入0.2g/LCoCl2,最终效价提高了约30%。发酵罐实验的结果显示,Co2+的加入对菌体的生长并没有促进作用,它能提高红霉素效价可能是由于它对菌体的代谢过程产生了影响。己糖激酶在整个发酵过程中的比活变化不大,基本上稳定在0.01U/mg。6-磷酸葡萄糖脱氢酶与磷酸果糖激酶的变化趋势基本上相反,在发酵前期(50h之前)前者比活较低,后者高,发酵中后期(80h之后)前者升高,后者降低。可以认为在葡萄糖总量一定时,细胞内HMP途径与EMP途径达到动态平衡,当一个途径的通量变大时,另一个途径的通量会相应减小。在红霉素生物合成中,由于需要琥珀酰CoA作为合成甲基丙二酰CoA的底物,因此适当加强EMP途径和TCA循环,并在发酵后期减弱HMP途径可能会对红霉素合成有利。6-磷酸葡萄糖脱氢酶比活在发酵中后期维持在较高水平可能是此时菌体粘度下降,不断补入氮源,导致菌体生长旺盛所致。结合有机酸含量的变化也可以看到,当柠檬酸含量不断降低时,α-酮戊二酸含量不降反升,说明α-酮戊二酸不仅来自TCA循环,还可能由于此时补入的有机氮源被菌体利用,氨基酸的脱氨转氨作用导致α-酮戊二酸含量增加。这需要进一步的实验证明。异柠檬酸脱氢酶的前一个高峰出现的时间与其他糖代谢酶一致,推测是由于菌体生长因而TCA循环较为活跃;而后一个高峰则与效价快速增长的时间相吻合。但是在发酵100h之后,对照中的比活开始不断升高,而加入Co2+的比活此时略有上升,但很快又有所下降。结合图1的效价变化可以看出,加入Co2+的效价增长率基本保持不变,而对照中在100h后效价的增长率略有降低,甚至出现负增长,推测在对照中三羧酸循环流量的增加并没有使红霉素合成前体(如琥珀酰CoA)增加,而可能转移到了其他途径。
甲基丙二酰CoA异构酶和甲基丙二酰CoA羧基转移酶对红霉素内酯环前体甲基丙二酰CoA的合成有重要作用。两者的比活高峰均出现在前期(50h左右),说明它们在菌体生长时就已经存在,除参与红霉素合成外,也参与菌体生长。发酵后期比活降低,如果能在此时提高这两个酶的比活也许可以促进红霉素的生产。Co2+的加入,对甲基丙二酰CoA异构酶的比活有非常明显的促进作用,而对甲基丙二酰CoA羧基转移酶也有一定影响。加Co2+的发酵液中有机酸含量普遍比对照高,所检测的有机酸都是参与TCA循环的,因而可以认为Co2+的加入能提高细胞中TCA循环的通量,增加红霉素合成前体琥珀酰CoA的含量,对提高红霉素产量有利。综上所述,在培养基中添加浓度为0.2g/L的CoCl2能显著提高红霉素发酵的水平,这可能是由于Co2+提高了甲基丙二酰CoA异构酶和甲基丙二酰CoA羧基转移酶的比活,使得合成大环的前体增多,从而提高红霉素生产水平。另一方面,添加Co2+能一定程度上提高糖代谢各关键酶的比活,使糖耗相应加快,促进TCA循环,也有利于红霉素生产。由于微生物的次级代谢非常复杂,本文只是对红霉素生物合成过程的糖代谢及次级代谢做了初步研究。关于整个代谢过程更深入的了解(如氮源代谢过程等)将有助于对初级代谢与次级代谢之间的关系有深刻的认识,从而指导实际生产。
相关热词搜索:红霉素
上一篇:利用原生质体融合技术选育高产菌株
下一篇:大孔树脂在红霉素提取中的应用进展
分享到:
 收藏
收藏
收藏
评论排行
- ·中国发酵企业数据库(4)
- ·(4)
- ·CoQ10高产菌株选育的研究进展(2)
- ·抗生素发酵工艺所用冷却塔的性能分析及处理(1)
- ·微生物菌种选育技术.rar(1)
- ·发酵生产染菌及其防治(1)
- ·赤藓糖醇发酵工艺研究(1)
- ·重组AiiA 蛋白可溶性表达及发酵条件优化(1)
- ·生物反应器设计软件_发酵罐绿色版(1)
- ·酵母粉、酵母浸粉的区别(1)
- ·雷帕霉素研究进展(1)
- ·透明质酸用途和行业概况(1)
- ·黄酒制作工艺(1)
- ·水解(酸化)工艺与厌氧发酵的区别(1)
- ·糖蜜酒精废液处理过程中产生的微生物蛋...(1)
- ·紫杉醇高产菌发酵产物的分离、纯化和鉴定(1)
