酿酒酵母非发酵代谢调控
2007-06-17 14:48:04 来源:网络数据库 评论:0 点击:
Although sugars are clearly the preferred carbon sources of the yeast Saccharomyces cerevisiae, nonfermentable substrates such as ethanol, glycerol, lactate, acetate or oleate can also be used for the generation
of energy
ies of the peroxisomal enzymes involved in b-oxidation increase upon growth with oleate [intensively investigated for the b-keto thiolase gene FOX3 (POT1) and the catalase A gene CTA1]. Transfer of yeast cells from glucose-containing medium to glycerol or ethanol leads to a substantial (2- to 10-fold) derepression of the corresponding genes. In the presence of oleate, gene expression is further induced (2- to 20-fold; Einerhand et al. 1991; Filipits et al. 1993; Karpichev and Small 1998). Both derepression and induction of these genes require a functional Snf1 protein kinase (Simon et al. 1992; Filipits et al. 1993). In contrast, deletion of MIG1 did not alter the regulation of FOX3 (Einerhand et al. 1995). However, FOX3 and some functionally related genes are repressed by Ume6, which binds to URS1 sequence motifs. Interestingly, peroxisomal functions are also affected by retrograde regulation and RTG genes. Loss of mitochondrial DNA (q0) not only leads to an increased transcription of the CIT2 gene encoding the peroxisomal citrate synthase but also stimulates the expression of POX1, FOX2, CTA1,SPS19 and PEX11 (Chelstowska and Butow 1995;Epstein et al. 2001). Consequently, functional RTG genes are necessary for the efficient utilization of oleate. While these data indicate that functional RTG genes may be required for efficient oleate induction, other results argue for a promoter strength which is almost unaffected by a rtg1 mutation (Kos et al. 1995).
Derepression of genes encoding peroxisomal proteins is mediated by Adr1-binding sites (shown for genes POX1, FOX2, FOX3, CTA1, SPS19, PEX1, PEX11;Filipits et al. 1993; Cheng et al. 1994; Simon et al. 1995;Gurvitz et al. 2000, 2001). The transcript level of CTA1,FOX2, FOX3 and PEX1 was clearly reduced in an adr1 mutant, while overexpression of ADR1 could substantially relieve glucose repression of CTA1 transcription (Simon et al. 1991, 1995).
Gene induction by oleate is mediated by a distinct promoter motif, designated oleate response element (ORE; Einerhand et al. 1993; Filipits et al. 1993). From the comparison of sequence variants upstream of boxidation genes, the 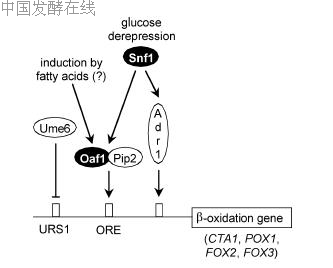
相关热词搜索:Gene regulation Transcriptional acti
上一篇:毕赤酵母表达系统
下一篇:微生物发酵处理对豆粕抗营养因子的影响
分享到:
 收藏
收藏
收藏
评论排行
- ·中国发酵企业数据库(4)
- ·(4)
- ·CoQ10高产菌株选育的研究进展(2)
- ·抗生素发酵工艺所用冷却塔的性能分析及处理(1)
- ·微生物菌种选育技术.rar(1)
- ·发酵生产染菌及其防治(1)
- ·赤藓糖醇发酵工艺研究(1)
- ·重组AiiA 蛋白可溶性表达及发酵条件优化(1)
- ·生物反应器设计软件_发酵罐绿色版(1)
- ·酵母粉、酵母浸粉的区别(1)
- ·雷帕霉素研究进展(1)
- ·透明质酸用途和行业概况(1)
- ·黄酒制作工艺(1)
- ·水解(酸化)工艺与厌氧发酵的区别(1)
- ·糖蜜酒精废液处理过程中产生的微生物蛋...(1)
- ·紫杉醇高产菌发酵产物的分离、纯化和鉴定(1)
