阿维菌素发酵过程有机酸积累规律与生物合成的关系
2007-11-05 09:07:15 来源:华东理工大学学报 评论:0 点击:
微生物细胞体内有众多的代谢反应,这些代谢反应组成了复杂的代谢网络并受到严格调控。由于有相当数量的代谢反应,尤其是一些糖代谢的关键节点都与有机酸的代谢密切相关,所以可通过检测体系内有机酸的积累情况来研究物质在各个途径上的分布情况,最终更加深入理解细胞的代谢调控机制[1~4]。阿维菌素糖苷配基的合成是通过7个乙酸单位和5个丙酸单位头尾缩合而成[5],这些有机酸单位的直接来源是乙酰CoA,所以阿维菌素的合成与糖代谢有着密切关系。在葡萄糖的代谢途径中间有机酸是非常重要的中间产物[6],如丙酮酸是糖酵解途径的终点产物,它可以转化为乙酰CoA;而柠檬酸、α-酮戊二酸、琥珀酸则是柠檬酸循环的中间体。
为了进一步了解阿维菌素发酵过程的特性,测定了实际生产过程中几个不同发酵批次4种有机酸在胞内与胞外的积累情况,并与产素速率进行关联性分析,结果发现高产批次与低产批次有机酸的代谢情况差异明显。
1 材料与方法
1.1 菌种与培养基
菌种:研究中用到的Streptomyces avermitilis突变株Av7为浙江海正药业提供。发酵培养基(g/L):淀粉50,热榨黄豆饼粉5,花生饼粉14,酵母粉4,酵母膏1,玉米浆1,CoCl2·H2O 0.03,pH 7.0自来水配制。
1.2 有机酸的测定
1.2.1 流动相的配制 流动相优化实验表明,样品中各组分在磷酸溶液的pH为2.32时分离度最高。
取1.15 mLw=0.85的磷酸,用超纯水稀释并配制成0.01 mol/L的磷酸缓冲液,用NaH2PO4调节pH至2.32,0.45μm合成纤维素酯膜进行真空超滤,超声波脱气后备用。
1.2.2 标准品的配制 精确吸取0.38 mL乙酸,
称取0.4 g丙酮酸、柠檬酸、琥珀酸加入1 000 mL容量瓶中,用超纯水溶解并配成0.4 g/L的有机酸混合标准储液,然后进行稀释,分别配制成浓度为0.05、0.5、1.0、1.5、2.0、3.0 g/L的系列有机酸标准溶液。以上有机酸均为生化试剂。
1.2.3 发酵样品的HPLC分析 取阿维菌素发酵
的发酵液30 mL,离心去除菌体,将滤液冷冻保存备用。分析时取1 mL滤液16 000 r/min离心10 min,上清液0.22μm的膜过滤后进样分析。菌体用预冷至4°C的生理盐水清洗2遍,再浸没于去离子水中,稀释到原发酵液体积,超破壁,12 000 r/min4°C条件下离心分离,上清液冷冻保存备用。胞内样品测定预处理方法同前。
1.2.4 色谱条件 色谱系统:Anglent1100。色谱柱:AquaSep.公司C8(5μm,4.6 mm×25 cm)。流动相:0.01 mol/L磷酸水溶液(用NaH2PO4调节至pH2.32);流速:0.6 mL/min。进样量:20μL。柱温:30°C。检测波长:210 nm。各种有机酸出峰时间(min)依次为:丙酮酸6.536;α-酮戊二酸7. 728;乙酸8. 432;柠檬酸9.644;琥珀酸11.168。发酵液中由于有其他氨基酸或者杂质的干扰,各种有机酸的出峰时间与标准的相比有一定偏差。
1.3 测定方法
效价测定:取发酵液3 mL,加入无水乙醇7mL,充分振荡5 min,静置4 h,过滤取滤液采用HPLC法测定阿维菌素B1a的效价。色谱条件:Anglent1100型高效液相色谱仪:C18反相柱(250mm×4.0 mm);流动相为V(甲醇)∶V(水)=9∶1;流速为1.0 mL/min;检测波长为246 nm;柱温25°C;进样量20μL;B1a标准品由浙江海正药业提供。
总糖与还原糖测定使用菲林法[7]。固形物含量PMV(Packed Mass Volume)采用离心法计算,取10 mL发酵液,3000 r/min离心15 min,计算固形物体积占发酵液的比例。
1.4 数据分析
分别跟踪了一个高产批次和一个低产批次的胞内胞外有机酸浓度与效价的变化规律。效价与发酵液体积经过换算可以得到发酵产物总量。以此数据使用差商方法计算得到各个时间点上的产素速率。进一步分析表明有机酸浓度变化与产素速率之间存在一定的关联。
2 结果与讨论
2.1 高产批次与低产批次基本发酵参数的比较在比较高产批次与低产批次的离线数据时发现它们前期离线参数的变化非常类似。如图1所示,80h前总糖浓度的变化基本重合,也就是碳源的利用速率基本一致,其他还原糖、PMV等的过程曲线也有类似现象。真正体现出差异的除了产素能力以外,就是pH值,低产批次一直要低于高产批次。我们进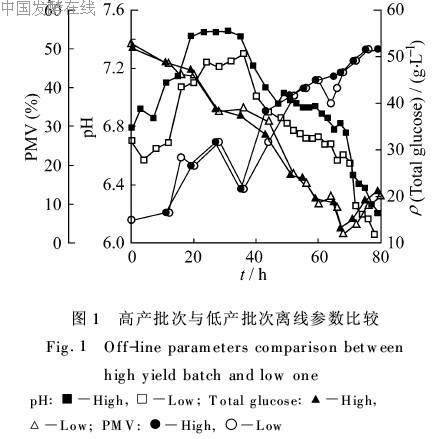
行的其他实验结果证明(未列出),发酵过程的前期对于阿维菌素的合成相当重要,前期生长的差异会导致菌丝形态分化的异常,代谢流量分布的改变以及产素能力的变化。从观察到的现象看来,前期的主要不同体现在H+平衡上,因此本文将进一步考察有机酸浓度变化的规律。
2.2 高产批次初期丙酮酸在胞外积累
丙酮酸的分布基本倾向于输出胞外,胞外浓度往往是胞内的数倍。高产批次中胞外的丙酮酸浓度在初期保持较高水平,40 h开始下降,至80 h后稳定在初期的25%水平,如图2(a)。低产批次则明显不同,初期胞内丙酮酸浓度甚至高于胞外,但是进入产素期后反而出现大量排出胞外的现象。
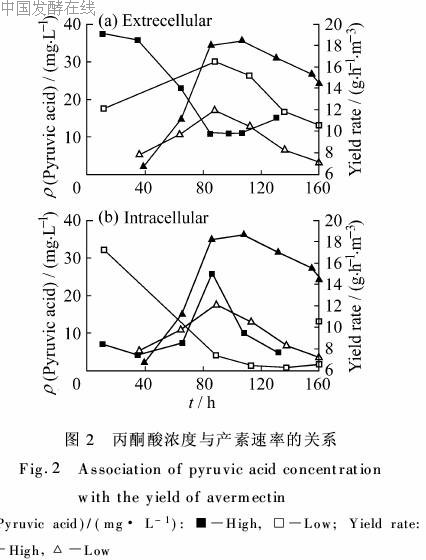
从多个批次的情况看来,较好的批次胞外丙酮酸的初期水平较高,但实际胞内浓度却相对较低,这表明早期的初级代谢是受到限制的。分析发酵过程尾气数据时也发现初期CER、OUR低于20 mo(m3·h)的批次最后产素情况较为理想,可以作为另外一方面的证据。产素期低产批次胞内浓度远低于胞外,比较高产批次可以发现后者除了在60~100 h处胞内浓度出现峰值,后面基本与外部浓度相当。有文献提出添加丙酮酸抑制产素,但从本实验结果来看丙酮酸并非越少越好。从碳源的代谢途径看来,丙酮酸主要作为乙酰CoA的前体,乙酰Co既可流入TCA循环产生能量,也可作为阿维菌素糖苷配基拼装的必要前体。所以在一般阿维菌素的发酵中间胞外丙酮酸的高浓度更多的是一种结果而非产素速率下降的主要原因。如高产批次的丙酮酸胞内浓度要高于低产批次,体现出代谢途径的活跃,也能确保产素的前体供应。低产批次却在产素期出现胞内浓度极低,胞外发生丙酮酸积累,这本身就说明丙酮酸的利用出现问题,流向产素的前体原料有限,自然产素速率低。
2.3 乙酸浓度与产素的关系
乙酸是阿维菌素合成的可能前体之一,也能参加初级代谢。高产批次中间,乙酸在160 h前一直在被摄入胞内,胞内胞外的浓度之比从11 h的0.34一路升至131 h的7.74。胞内乙酸浓度较高的时间正是产素速率较高的阶段,说明两者存在一定关联(图3)。
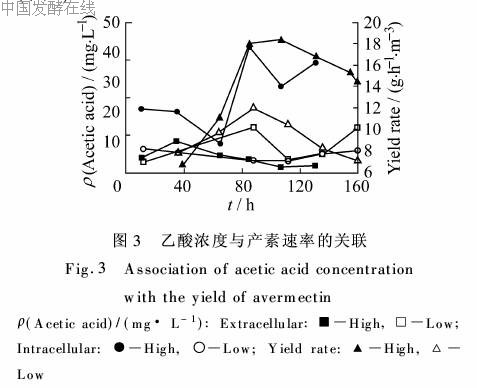
胞内很可能存在一种机制,就是将暂且不用的营养物质释放到胞外,在生长阶段,乙酸盐并非最好的碳源物质,自然就在胞外形成积累;而当产素速率加快,前体供应紧张的时候,又必然地需要摄入以满足需求。低产批次直到160 h依然是大量乙酸盐处于胞外,胞内浓度较低,说明此类前体并非此时制约产素的因素。
2.4 柠檬酸和琥珀酸不同批次的分布差异
柠檬酸与琥珀酸源自TCA循环,是糖代谢的中间产物。高产批次两种酸都是在80 h之前胞外出现积累,此后胞外浓度一直较低,而胞内浓度则一直保持较高水平。低产批次主要在胞外积累,胞内浓度一直较低,说明此时生成的有机酸已经超过需求所以被释放到环境中(图4)。由于链霉菌中琥珀酰CoA也可能再异构化成2-甲基丙二酰CoA[8],后者是阿维菌素糖苷配基的合成启动单元,对产素意义重大。因此胞内维持这两种有机酸在较高浓度可能对阿维菌素的生物合成有利。
3 结 论
生产中的罐批,初始条件基本一致,但最后的产素结果却可能差异较大。比较离线参数有时会发现参数变化相当近似。例如上面分析中发现实际出现差异的就是细胞内外的有机酸积累情况。某种意义上来说,胞内的有机酸浓度体现的是相关途径的活跃程度,胞外积累的有机酸则是暂时因为胞内供过于求而排出的部分。在综合分析了多种有机酸浓度与产素速率关系后,可以发现在低产批次中存在一个共性,那就是尽管胞内有机酸浓度较低,但是胞外在80 h后都出现积累。究其原因,可能是生长期延长以致与产素期重叠过多造成。
低产批次往往初期菌体活力非常高,生长迅速,消耗中间产物也快,所以不会形成积累。但是如不采取有效措施控制,生长阶段与产素阶段重叠,依然大量产生有机酸,在代谢流量的分配上却偏向生长,但是菌量却因其他的因素而接近极限,自然就出现了胞内代谢供过于求和胞外的有机酸积累。这也就是为什么前面一些批次中虽然补料速率和糖耗都较高,但是产素速率却偏低的原因。
相关热词搜索:阿维菌素
分享到:
 收藏
收藏
收藏
评论排行
- ·中国发酵企业数据库(4)
- ·(4)
- ·CoQ10高产菌株选育的研究进展(2)
- ·抗生素发酵工艺所用冷却塔的性能分析及处理(1)
- ·微生物菌种选育技术.rar(1)
- ·发酵生产染菌及其防治(1)
- ·赤藓糖醇发酵工艺研究(1)
- ·重组AiiA 蛋白可溶性表达及发酵条件优化(1)
- ·生物反应器设计软件_发酵罐绿色版(1)
- ·酵母粉、酵母浸粉的区别(1)
- ·雷帕霉素研究进展(1)
- ·透明质酸用途和行业概况(1)
- ·黄酒制作工艺(1)
- ·水解(酸化)工艺与厌氧发酵的区别(1)
- ·糖蜜酒精废液处理过程中产生的微生物蛋...(1)
- ·紫杉醇高产菌发酵产物的分离、纯化和鉴定(1)
