表达胰岛素的毕赤酵母生长动力学及诱导策略
2007-09-16 23:11:00 来源:北京化工大学学报 评论:0 点击:
巴斯德毕赤酵母(Pichia Pastoris)自上世纪80年代被开发为外源蛋白表达系统以来一直被广泛应用,一些重组蛋白已成为正式产品进入了市场[1]与其他真核表达系统相比,巴斯德毕赤酵母有一个显著的优势,即容易实现高密度发酵而且外源蛋白分泌效率高[2]。另外,巴斯德毕赤酵母生存能力强,即使在廉价的基础盐培养基中也能实现高密度发酵,可进行大规模生产。胰岛素是治疗糖尿病的常用特效药物,有很大的市场需求,近年来重组人胰岛素已占领了市场的大部分份额[3]。对于分泌性表达的蛋白,细胞密度越高意味着表达水平越高[4],所以高密度培养是毕赤酵母高效表达外源蛋白的重要因素之一。本文对实验室构建的重组人慢速胰岛素基因的毕赤酵母因工程菌生长动力学进行研究,以探究其是否具有高密度发酵的潜力;在优化了的基础盐培养基上,为有效提高工
程菌表达目的蛋白的产量及产物对细胞的得率,研究了混合诱导碳源以及在发酵液中添加其它混合物对目的蛋白产量的影响,结果使目的蛋白产量提高了将近一倍。10L发酵罐发酵试验得目的蛋白产量是摇瓶发酵水平的5倍多,为进一步放大培养和指导工业化提供参考。
1 材料与方法
1·1 材料
1·1·1 菌种 重组毕赤酵母基因工程菌由本实验室构建并保存。
1·1·2 培养基 种子培养基(YPG)由1 g酵母提取物、2 g大豆蛋白胨、3 mL甘油溶于100 mL去离子水中制成;基础盐培养基以及微量元素配方参照文献[5]。
1·1·3 仪器 DYY-Ⅲ5型稳压稳流电泳仪(北京六一仪器厂);WFB-UV-2000型紫外可见分光光度计 (龙尼柯仪器有限公司,上海);HZQ-X100振荡培养箱 (哈尔滨市东联电子技术开发有限公司);GUJS-10C型机械搅拌发酵罐 (镇江东方生物工程设备技术有限责任公司)。
1·2 方法
1·2·1 摇瓶培养 新鲜平板上挑取单菌落接到yPG培养基中,并在30℃和200 r/min的条件下振荡培养过夜。然后,以10%的接种量转接到基础盐培养基中,在30℃和200r/min的条件下振荡培养。其间取样检测甘油浓度,待甘油耗尽,饥饿1 h开始诱导。诱导期每12 h补加甲醇进行诱导。诱导期还加入混合碳源甘油(甘油与甲醇体积比为1∶6)或山梨醇(1M山梨醇溶液与甲醇体积比为1∶6)进行试验,并在此试验基础上加入混合物组氨酸、天门冬
氨酸、酵母提取物、大豆蛋白胨以及微量元素进行试验。诱导96h时停止发酵。
1·2·2 发酵罐发酵 新鲜平板上挑取单菌落接到50mL YPG培养基中,并在30℃和200 r/min的条件下振荡培养过夜,然后接入2级种子450mL YPG培养基中,继续培养24 h,再以10%的接种量转接到装5L基础盐培养基的10L发酵罐中进行批次补料培养,培养温度为30℃,pH控制在5·5~6·0之间,通过调节通气量(3~12L/min)和搅拌转速(200~500r/min)使溶解氧体积分数保持在30%左右。当甘油耗尽时每小时补加甘油50mL,补加4 h。当补加的甘油耗尽时,饥饿1 h开始补加甲醇(补加的甲醇中含4mL/L的微量元素)进行诱导。在诱导24h后加入酵母提取物、大豆蛋白胨以补充养分,诱导96h下罐。
1·2·3 分析方法 将菌液稀释后于波长600nm处以去离子水为参比,用比色法测定细胞吸光度;细胞质量浓度测定参照文献[6],测定所取发酵液中的细胞干质量,再除以所取发酵液体积即得细胞质量浓度;甘油浓度用高碘酸钠氧化法测定[7];发酵液中总蛋白浓度测定参考考马斯亮蓝染色法[8];发酵液中目的蛋白(慢速型人胰岛素原)浓度的测定法是取发酵液离心后的上清走SDS-PAGE,将凝胶进行计算机扫描分析,用BIO-PRINT凝胶成像与分析
系统分析目的蛋白的含量,根据考马斯亮蓝染色法测定的发酵液中总蛋白的量推算出目的蛋白浓度;发酵动力学参数分析计算方法参照文献[9]。
2 结果与讨论
2·1 基因工程菌摇瓶培养生长阶段
基因工程菌由于其被整合的外源蛋白基因的不同,外源蛋白基因整合到宿主染色体位置的不同和外源基因载体的构建方式的不同,其生长规律有可能不尽相同。将本实验室构建的基因工程菌毕赤酵母所测细胞吸光度(A600)与细胞质量浓度按最小二乘法进行线性回归,得到细胞质量浓度与A600呈线性关系,回归方程为CS=0·2303A600-0·1101(方差R2=0·9995),式中,CS为细胞质量浓度,g/L。所以在以下的试验中利用测得的发酵液A600就可以推算出发酵液中的细胞质量浓度。将种子液接入基础盐培养基,每2 h取样测细胞光密度以及甘油残留浓度。试验结果列于表1。
2·1·1 基因工程菌生长模型的建立
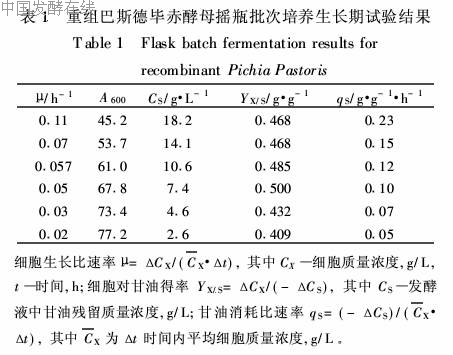
1
μmax,式中,μmax为细胞最大生长比速率,Ks为饱和常数。以1/μ对1/CS作图,如图1所示。结果1/μ与1/CS呈线性关系(R2=0·9848),表明在其他底物相对过剩时本基因工程菌的生长与限制性基质甘油残留浓度的关系符合Monod关系式,将1/μ与1/CS按最小二乘法进行线性回归得μmax=0·204h-1,Ks=24·3g/L。
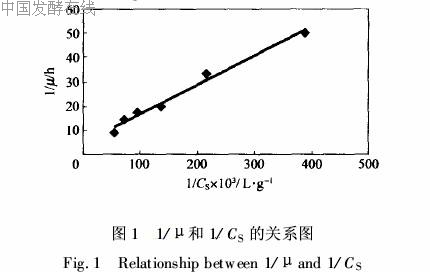
2·1·2 理论最大细胞对甘油得率Y*X/S和菌体生长维持系数m的推导 对于在热力学上远离平衡态的活细胞,为了维持其生命,必须要获取高能物质并将其化学能转变为热能,用以维持其渗透压,修复DNA、RNA和其他大分子,因此能量不仅要消耗于细胞的生长上,而且也要消耗在维持细胞结构上。对基质消耗的物料衡算方程中必须考虑维持代谢这一项[9],qS=1Y*X/Sμ+m。其中Y*X/S为生成细胞(干质量)与完全消耗于细胞生长的甘油的质量比,它表示了在无维持代谢时的细胞得率,可称为理论最大细胞对甘油得率;m为菌体生长维持系数。在微生物生长阶段用于维持的碳源消耗量大致恒定,且与生长无关。通过qS对μ进行最小二乘法线性
回归,如图2所示,得Y*X/S=0·497 g/g,m=0·007
g/g·h (R2=0·996)。从表1可以看出,试验测得细胞对甘油得率随生长比速率的减小而呈现先增大后减小的趋势,甚至大于理论最大细胞对甘油得率。在细胞生长阶段后期补加甘油可以有效阻止细胞对甘油得率减少的趋势;用于维持细胞结构的菌体生长维持系数m=0·007g/g·h,相比大肠杆菌利用葡萄糖作为底物生长时的细胞维持系数0·057 g/g·h[10]小得多,这说明细胞利用甘油主要是用来生长,只有很少一部分用来维持细胞结构。由于维持细胞结构的碳源是非生产碳源,所以m的大小可作为评价菌株优劣的指标之一,m值越小,说明菌株高密度培养潜力越大。因此本株菌相比大肠杆菌高密度培养的优势明显,就细胞高密度培养来说是一株优良菌,具有高密度培养的潜力,可用来作为工业化生产的工程菌。
2·2 基因工程菌摇瓶培养诱导阶段
2·2·1 诱导期辅助碳源的选择对产物产量的影响
毕赤酵母甲醇诱导时流加其他碳源,可促进细胞的能量代谢和细胞的分解与合成代谢的强度,有利于表达量的提高[11]。通常所使用的碳源甘油对产物的表达有抑制作用。而山梨醇对甲醇氧化酶启动子AOX1没有阻遏作用,据文献报道[11]用它作为甲醇诱导的辅助碳源对外源蛋白的表达有利。经过甘油为底物的生长期后,分别利用甲醇、甲醇和甘油的混合碳源以及甲醇和山梨醇的混合碳源进行诱导,保持发酵过程中操作和其他参数基本一致的情况下比较这三种条件的优劣。每隔12 h在进行补料的同时取样测细胞光密度以及目的蛋白浓度。诱导期发酵液中细胞浓度与目的蛋白浓度变化如图3和图4所示。相关数据列于表2。
在利用甲醇为唯一碳源诱导时,目的蛋白产量为26mg/L,诱导结束时的细胞质量浓度为21·2g/L,菌体的生长比速率μ、目的蛋白对细胞得率YP/X和目的蛋白生成比速率qP分别为0·005 h-1、1·2mg/g和0·013mg/g·h。在利用甘油作为甲醇诱导的辅助碳源时,虽然细胞质量浓度和μ有所提高,但是目的蛋白产量以及YP/X和qP都明显下降,说明甘油的加入虽然有助于提高细胞质量浓度,但是对产物生成的抑制作用明显,不适合用来作为甲醇
的辅助碳源。
在利用山梨醇作为甲醇诱导的辅助碳源时,细胞质量浓度和μ也比甲醇作为碳源时提高了,但是发酵液中目的蛋白的浓度在诱导48 h左右达到最大值,然后开始下降或者不再增长,表明诱导后期山梨醇对本株菌产物的分泌起抑制作用。诱导结束时目的蛋白产量略低于单一甲醇碳源诱导水平,而YP/X和qP分别为单一甲醇碳源诱导水平的75%和61%。综上所述选择甲醇为唯一碳源进行诱导效果最佳。
2·2·2 诱导期添加混合物对产物表达的影响 根据2·2·1试验结果,在以甲醇为碳源进行诱导的基础上,研究诱导时流加其他混合物对产物的影响。本实验室所设计的慢速型人胰岛素原分子中,组氨酸和天门冬氨酸所占比例最大,分别占蛋白分子量的11%左右,所以在诱导期毕赤酵母进行的合成产物代谢中组氨酸和天门冬氨酸的缺乏有可能成为阻碍蛋白合成的因素之一,称为瓶颈效应。试验中诱导期每升发酵液中添加2·5 g的组氨酸和2·5 g的天门冬氨酸,结果诱导结束时目的蛋白产量为36mg/L,是单一甲醇诱导的1·4倍,YP/X和qP都超过单一甲醇诱导水平。另外,酵母提取物和大豆蛋白胨里也富含多肽和氨基酸以及少量的维他命和微量元素,而这些组分在诱导期能提高细胞得率和合成分泌外源蛋白的能量[12]。试验中诱导期每升发酵液中添加2·5g的酵母提取物和5g的大豆蛋白胨,在诱导结束时,目的蛋白产量达到49 mg/L,几乎比单一甲醇诱导增长一倍,而且YP/X和qP分别是单一甲醇诱导时的1·7倍和1·6倍。与发酵液中添加组氨酸和天门冬氨酸的试验数据YP/X和qP相比,细胞产目的蛋白的效率更高,效果尤胜。考虑到酵母提取物和大豆蛋白胨比组氨酸和天门冬氨酸廉价易得,在发酵过
程中可以用酵母提取物和大豆蛋白胨代替组氨酸和天门冬氨酸作为诱导时的添加物。当诱导阶段每升发酵液中添加1 mL的微量元素时,诱导结束时目的蛋白产量为44mg/L,YP/X为1·6mg/g,并且细胞质量浓度也达到27·5g/L,较之单一甲醇诱导时都有增加。所以诱导阶段补加微量元素是非常有必要的。
2·3 基因工程菌的发酵罐培养
根据摇瓶发酵试验结果,将基因工程菌进行上罐培养。诱导阶段在流加的甲醇中加入4 mL/L的微量元素,并且在诱导24 h后加入酵母提取物、大豆蛋白胨(加入量与摇瓶培养试验相同)以补充养分。由于发酵罐设备的限制,在菌体生长高峰期和分泌蛋白高峰期无法满足细胞对氧的需求,而且发酵罐中溶解氧体积分数一度降到接近0%,影响了重组毕赤酵母的高密度发酵和分泌目的蛋白产量。发酵120h下罐时,细胞质量浓度为57·5g/L,目的蛋白产量达到256mg/L,蛋白产量是摇瓶发酵蛋白产量的5倍多。由于发酵罐中氧的供应好于摇瓶培养,所以诱导期YP/X和qP较之摇瓶培养都大大提高。作者相信如果解决了发酵中氧的供应阶段性不足的缺陷,细胞质量浓度和目的蛋白产量还会增高。
3 结论
工程菌发酵生长阶段菌体的生长与限制性基质甘油残留浓度的关系符合Monod关系式,通过1/μ对1/CS进行线性回归得μmax=0·204 h-1,Ks=24·3g/L。通过qS对μ进行线性回归,得理论最大细胞对甘油得率Y*X/S=0·497 g/g以及菌体生长维持系数m=0·007g/g·h,后者说明本株菌就细胞高密度培养来说是一株优良菌,具有高密度培养的潜力。利用甘油或者山梨醇作为甲醇诱导的辅助碳源效果不如单一甲醇诱导,诱导期补加酵母提取物、大豆蛋白胨和微量元素能有效的提高目的蛋白产量。10L发酵罐上罐试验120 h,细胞终质量浓度为57·5g/L,目的蛋白产量为256mg/L,蛋白产量是摇瓶发酵蛋白产量的5倍多。
相关热词搜索:胰岛素
上一篇:应用响应面法优化武夷菌素发酵培养基
下一篇:异亮氨酸对他克莫司发酵的影响
分享到:
 收藏
收藏
收藏
评论排行
- ·中国发酵企业数据库(4)
- ·(4)
- ·CoQ10高产菌株选育的研究进展(2)
- ·抗生素发酵工艺所用冷却塔的性能分析及处理(1)
- ·微生物菌种选育技术.rar(1)
- ·发酵生产染菌及其防治(1)
- ·赤藓糖醇发酵工艺研究(1)
- ·重组AiiA 蛋白可溶性表达及发酵条件优化(1)
- ·生物反应器设计软件_发酵罐绿色版(1)
- ·酵母粉、酵母浸粉的区别(1)
- ·雷帕霉素研究进展(1)
- ·透明质酸用途和行业概况(1)
- ·黄酒制作工艺(1)
- ·水解(酸化)工艺与厌氧发酵的区别(1)
- ·糖蜜酒精废液处理过程中产生的微生物蛋...(1)
- ·紫杉醇高产菌发酵产物的分离、纯化和鉴定(1)
