白腐真菌漆酶的研究进展及应用前景
2010-10-23 09:38:33 来源:南华大学 药学与生命科学学院 湖南衡阳 421001 评论:0 点击:
漆酶是一种含铜的多酚氧化酶,与抗坏血酸氧化酶和哺乳动物血浆铜蓝蛋白同源,都属于蓝色多铜氧化酶家族。担子菌亚门的白腐真菌是分泌漆酶的主要真菌。文章主要对白腐真菌漆酶的特性、作用机理、研究概况以及在生产中的应用状况等方面进行了阐述,并提出了应用前景及发展趋势
摘 要:漆酶是一种含铜的多酚氧化酶,与抗坏血酸氧化酶和哺乳动物血浆铜蓝蛋白同源,都属于蓝色多铜氧化酶家族。担子菌亚门的白腐真菌是分泌漆酶的主要真菌。文章主要对白腐真菌漆酶的特性、作用机理、研究概况以及在生产中的应用状况等方面进行了阐述,并提出了应用前景及发展趋势。
关键词:白腐真菌漆酶 特性 催化机理 研究进展 应用前景
木质素是以苯丙烷类似物为单元构成的一大类生物多聚体,组成单元的结构及其连接键复杂而稳定。这种无规则的复杂结构使得木质素难以降解。自然界中木质素降解主要是通过丝状真菌,尤其是通过白腐真菌的分解作用来完成。白腐真菌降解木质素依赖一些酶的产生和分泌。这些酶共同构成了木质素降解酶系。白腐真菌分泌的木质素降解酶主要有木质素过氧化物酶、锰过氧化物酶和漆酶3种。
1 漆酶的简介
漆酶(氧化还原酶,EC
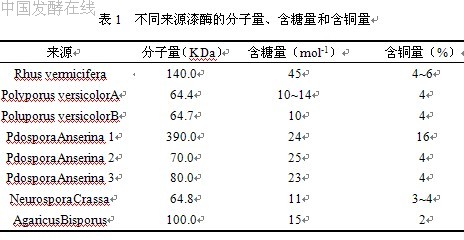
分泌漆酶的真菌主要分布于担子菌(Basidiomycoti-na)、多孔菌(Polyporus)、子囊菌(A scomycotina)、脉孢菌(Neurospra)、柄孢壳菌(Podospora)和曲霉菌(A spergillus)等属种,其中最主要的是担子菌亚门的白腐真菌。近年来,研究发现漆酶也存在于薰衣草链霉菌( Streptomycetes lavendulae )、灰色链霉菌(S·griseus)、苜蓿中华根瘤菌(Sinorhizobium meliloti)、Bacillus subtilis和Marinomonas mediterranea原核生物中。
据统计,漆酶能催化氧化六大类250余种底物,包括酚及其衍生物、芳胺及其衍生物、羧酸及其衍生物、金属化合物和其他非酚类底物等。一般,铜螯合剂对漆酶活性都有抑制作用,但是EDTA能增加P. conchatus漆酶的反应活性。不同的抑制剂对漆酶活性的抑制作用不同,如DEDTC(二乙基二硫代氨基甲酸盐)和巯基乙醇对P.spinctri-nus漆酶活性有强烈的抑制作用,而羟胺、EDTA对其基本上无抑制作用。pH值对漆酶活性也有一定的影响,大多数漆酶在碱性环境中不稳定、易失活。如何提高漆酶的稳定性已成为许多研究者研究高效漆酶的一个重要内容。有报道, C.thermophilium漆酶比一般的真菌漆酶具有更高的热稳定性,并且在pH值6.8时反应较适宜。
2 漆酶的研究进程
1883年日本吉田首次从漆树的汁液——生漆中发现漆酶,10年后Laborade等又证实真菌中也含有漆酶。现在认为漆酶是一种普遍分布于植物、昆虫、真菌和细菌中的重要酶种。20世纪末,漆酶受到广泛地关注。目前,一些大型真菌的产酶条件、漆酶基因的异源表达及其所分泌的漆酶的性质、特点是国内外学者研究的热点。
我国最早研究漆酶的是刘国智、黄葆同等,他们于20世纪50年代末利用漆酶在催化反应中需要消耗氧气的特性,设计了漆酶酶活的测定方法。但直到20世纪80年代末,国内对漆酶的研究一直停留在漆酶酶活的测定、漆酶表观特性的描述性研究上;与此同时,漆酶的分离纯化、动力学特性和固定化的研究也开始起步。近年来,随着分子生物学技术的不断发展,漆酶的研究也取得了令人瞩目的成绩,学者们已经测定了漆酶的氨基酸序列,克隆了它的基因,并研究了漆酶的编码基因及其表达调控机制。目前,漆酶的理论与应用研究已在生物、化学、物理、医学等多领域,分子、细胞、生物组织等多水平展开。漆酶在食品和环保工业上的应用研究也日益活跃。
3 白腐真菌漆酶的特性
漆酶能有效地降解木质素,它的结构决定了漆酶的特性。主要表现为:
3.1 氧化特性
不同的漆酶其氧化能力与方式也不相同。漆酶氧化速率的大小不仅与漆酶本身的稳定性有关,而且也与漆酶与底物作用的一些动力学参数有关。
3.2 对抑制剂的敏感性
一般来说,不同的抑制剂对漆酶活性的抑制作用不一样,同一种抑制剂对不同的菌种其抑制作用也不一样,抑制剂对漆酶活性的影响随抑制剂浓度的不同而不同,抑制剂浓度越大,其抑制作用越强。
3.3 底物专一性
漆酶催化氧化不同类型的底物已达250个,根据底物结构不同可分为酚类及其衍生物类、芳胺及其衍生物类、羧酸及其衍生物类、幽体激素和生物色素、金属有机化合物、其它酚类底物等六大类,并且还有增加的趋势,漆酶的这一性质预示着漆酶具有广泛的应用前景。
3.4 反应动力学
凡值的大小可以反映出漆酶对底物亲和力的大小,而漆酶对底物的亲和力又是影响漆酶氧化底物速率的一个重要因素。凡值越小,亲和力越大,漆酶对底物的氧化率也就越高。在一定的温度范围内,随着反应温度的升高,漆酶的酶活力增加。但是,温度越高,漆酶越不稳定,也就越易失活。pH值对漆酶酶活性亦有类似影响,一般来说,大多数漆酶在碱性环境中不稳定,易失活。漆酶反应的最适pH值随反应底物的不同而不同。
4 结构特征和催化机理
漆酶是一种多铜过氧化酶。通过对漆酶蛋白质晶体结构的研究,发现漆酶具有3个铜离子结合位点(T1、T2、T3),共结合4个铜离子(P.radiate漆酶除外,它仅含2个铜离子)。根据氧化还原电势、光学和磁学特征,划分为3种类型:
Ⅰ型铜离子具有典型的蓝铜谱带,紫外可见光谱上600nm[ε:5 000 mol/(L·cm)]处出现峰值,在电子顺磁共振光谱(EPR)在可见区显示4条电子转移条带,其中由Cys→CuⅠ产生的条带最强,是漆酶产生蓝色的主要原因;
Ⅱ型铜离子是由两分子的咪唑和一分子的水配位形成的松散的扭曲四面体结构,只具有一般的EPR谱带(A11>140×10
2个Ⅲ型铜离子分别与3个His中的N配位,同时与Tyr中的O配位形成氧桥组成双核铜区,具有抗磁性,因而在EPR上无谱带。铜离子是漆酶研究中的难点和热点之一。现已证实铜离子处于漆酶的活性部位,在漆酶的催化氧化过程中起着决定性作用。
漆酶催化反应可分为底物自由基中间体的产生和氧气还原成水2个方面。这2个方面都需要漆酶分子中的铜离子协同传递电子。首先,漆酶分子的Ⅰ型铜离子从还原态底物中吸收单个电子,底物被氧化成自由基,自由基引发各种各样的非酶促次级反应,如羟化、歧化和聚合等;其次,Ⅰ型铜离子将单个电子通过Cys-His途径传递给三核铜簇中心,从而氧被还原成水。铜离子处于漆酶的活性部位,在漆酶的催化氧化过程中起着决定性作用。其过程可
用公式表示为:
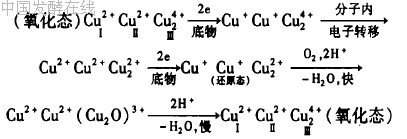
5 白腐真菌漆酶的分子生物学研究进展
5.1 漆酶基因的克隆
随着生物技术的发展,漆酶基因已经从木质素降解菌和非木质素降解菌中得到克隆,并且进行序列分析。许多来源的基因都是以家族形式存在于染色体上的。这些来源不同的基因表现出较高的同源性,目前已有近30种不同来源的漆酶基因被克隆和测序,包Coriolushir-sutus、Phlebia radiata、BasidiomycetePM1等。
1988年Frohman等首次利用快速扩增cDNA技术从白腐真菌(Pleurotus ostreatus)中克隆到漆酶poxI基因。后来,有学者采用RT-PCR技术获得漆酶基因。Giardina等运用该方法合成cDNA第一链和RT引物。引物可用来扩增一个750 bp cDNA片段。随后,很多学者利用建立的基因组文库或cDNA文库,然后设计探针在文库中筛选的方法来克隆漆酶基因。
5.2 漆酶基因结构和组织
真菌漆酶的基因外显子序列长度一般介于1 500~2 000 bp,整体的序列同源性并不高。漆酶基因中含有相当多的内含子,而且不同来源的漆酶基因内含子的位置大多是保守的。这2个方面可以作为分析酶的同源性和菌株间亲缘关系的依据之一。另外,异源真菌漆酶之间的氨基酸同源性较低,但在铜结合区具有较高的保守性。
5.3 漆酶基因的异源表达
对漆酶基因的克隆和序列分析都是为了实现漆酶的高表达。但多数漆酶自身的表达量很低,在异源宿主上表达重组漆酶蛋白是解决该问题的一个有效途径。
甲基营养酵母(Pichiapastoris)系统成功表达了若干种不同来源的漆酶,如Trame tessanguinea和Pycnoporus cinnabari-nus。Coprinus cinereus漆酶基因在Aspergillus oryzae中实现了漆酶的最好表达,表达产量达8~135 mg/L。Pycnoporuscinnabarinus漆酶Lacl基因在真菌宿主Aspergillus niger中实现了大量表达,表达量为70 mg/L。Coprinus cinereus漆酶基因和Pycnoporus cinnabarinus漆酶基因的异源高效表达,使得漆酶的大规模工业生产和应用成为可能。
6 漆酶的生产
漆酶在工业和生物技术领域已有广泛用途,有关漆酶生产方面的研究工作均围绕提高漆酶活性而展开。采用的方法是,筛选及选育优良的菌种,探索最适宜的培养方式及最佳生理培养条件,以使产漆酶菌株在工业中能得到高效应用。黄乾明等对白腐真菌(Trametes gallica)的孢子悬液用紫外灯诱变处理,筛选得到1株稳定高产漆酶的突变株,其峰值酶活力比出发菌株高4倍,达5002.6 U/L (ABTS法),且产酶稳定。
近十几年来,尤其是近五六年来,国际上有关这方面的研究取得了相当大的进展,然而仍存在着不少问题。
6.1 菌种的筛选及选育
生产漆酶的菌株多为真菌,近年来已经对漆酶产生菌进行了广泛的研究,但是选育漆酶高产菌株仍是当前的研究热点。随着分子生物学和基因工程的发展,构建产漆酶的工程菌株也逐渐成为研究的热点。
6.2 培养方式
目前研究最多的是液体培养,但也有采用固体培养的报道。这两种不同的培养方式对菌体生长和漆酶酶活的影响,现在还不是很清楚。有报道,Rigido-porus lignosus在固体培养条件下,漆酶和锰过氧化物酶的产量都较液体培养条件下大为提高,然而固体培养条件下营养物质、诱导剂、微量元素、温度及pH值等生理条件对漆酶的影响尚未见详细考察。在液体培养中,大多采用振荡培养,但是也有在静止培养条件下漆酶产量较高的报道。
6.3 培养基
目前,用于白腐真菌培养的培养基一般都是在kirk经典培养基的基础上改良的。漆酶的表达受培养基中碳源、氮源、碳氮比、金属离子和pH值等不同条件的影响。另外在培养基中加入某些天然物质,配成半合成培养基对提高漆酶的产量也非常有利。在合适的时间向培养基中加入适当浓度的诱导剂,也可以提高漆酶的产量;表面活性剂和某些金属离子对漆酶产量的提高也有一定的作用。
朱明旗等对亚香栓菌发酵产酶条件的研究结果表明,其产酶最适碳源为葡萄糖,最适氮源为硝酸铵,最适初始pH值6.0,最适培养温度28℃,5d时酶活达最大。
6.4 培养条件
在确定合适的培养基后,还要考察环境条件对菌体产漆酶的影响。需考虑的因素有菌龄、装液量、接种量、培养温度、培养时间、初始pH值、摇床转速及培养液黏度等。不同的菌种对这些条件的要求也不同。
6.5 分离纯化
漆酶的分离纯化时,一般先用快速定性滤纸过滤最优发酵条件下的发酵液,获得粗酶液,然后利用盐析、等电点沉淀、有机溶剂分级分离等方法对粗酶液进行处理;再用凝胶过滤、离子交换层析、吸附层析、亲和层析等方法进一步分离提纯,必要时选择区带电泳和等电聚焦作为最后的提纯步骤;最后还可以通过控制温度、加盐盐析、加有机溶剂或调节pH值等方法来获得漆酶晶体。
7 白腐真菌漆酶的应用前景
与化学氧化相比,由于生物氧化具有特异性、可降解性,以及反应条件的温和性等优点,使得漆酶在工业和生物技术领域具有广阔的应用前景。
7.1 农药的处理
20世纪,全球范围内农药的过度使用尤其是含氯的芳香类化合物,对环境造成了严重的污染。利用漆酶可以降解氯酚类有机化合物。T.versicolor漆酶对单氯酚进行24h处理后,邻位和对位氯酚的去除率分别达98.6%和100%。
7.2 污水的处理
在印染和造纸等工业过程中排放的大量含有染料的废水,成为当今主要的环境问题之一。采用通常的活性污泥方法处理该类废水很难达到预期目的。合成染料广泛应用于印染工业,目前已超过10 000种。根据染料结构的不同,可分为漆酶底物类染料和非漆酶底物类染料。蒽醌类染料是漆酶的底物,可被漆酶直接氧化,脱色和降解程度与酶活性呈正比。实验证明,白腐菌Trametes trogii(StrainBAFC463)分泌的漆酶和锰过氧化酶降解硝基苯和蒽醌混合物,12~24d后去除率大于90%。降解过程中漆酶的活性和稳定性远远高于锰过氧化酶,对污染物的降解起主导作用。偶氮类和靛青类染料不是漆酶的底物,但当添加小分子介体物质ABTS后,降解效果明显提高,提高比率与ABTS浓度呈正比。实验还证明,偶氮类和靛青类染料在漆酶作用下,降解非常缓慢,当添加(33 umol)蒽醌染料后,降解速度迅速提高到35~40 mg/(L·h),这一特点使漆酶在工业染料降解中的应用成为可能。
白腐真菌漆酶具有广泛的底物专一性,对纺织染料的脱色、降解效果明显。NOVO公司通过基因工程改造,用黑曲霉经深层发酵制得漆酶DeniliteⅡS,并且将其用于牛仔布靛蓝染料的脱色。
7.3 土壤的生物救治
Trine Eggen用含有Pleuoro-tus ostreatus的废弃培养基处理杂酚油污染过的土壤,在常温下培养7周,结果表明, 16种PAHs去除率达86%。Roper等还发现漆酶对稠环芳烃的去除能力与愈创木酚和2,6-二甲基苯酚有关。
7.4 造纸工业
应用漆酶进行纸浆的生物漂白是一个有潜力的研究课题。传统的氯气漂白工艺会产生有毒化合物而造成环境污染,而利用漆酶对纸浆进行漂白可以更有效地脱氯,不会产生有毒物质,以减少环境污染。
此外,漆酶能选择性地催化木质素降解,这一特性可用于纸浆生产。传统的纸浆是用烧碱或硫酸盐高温煮蒸原料,除去木质素制成的,这不仅会使得部分纤维素和半纤维素降解,影响纸浆得率,而且严重污染环境。用木质素分解酶进行木浆的前处理,是一种温和且清洁的脱木质化措施,而且还能够保证纤维素的完整。采用漆酶选择性降解木质素生产纸浆,能节约设备和能耗,缩短纸浆生产周期,降低生产成本,将会给造纸工业带来良好的经济效益和社会效益。还有,漆酶来源广泛、使用方便,这使漆酶介导系统已经得到了实际应用。
7.5 智能包装指示剂及生物检测
芬兰研究者利用漆酶的催化反应能形成有色产物的特性,研制出的渗漏指示剂可应用于智能包装方面。
漆酶在催化过程中消耗氧气,这一过程很容易被转化为电信号而被高灵敏地得到检测。利用这一原理可以制造各种类型的生物传感器。一种安培生物传感器用于测量酚类化合物已经运用于纸浆厂的污水测定。双酶系统传感器是漆酶在传感器研制中的新发展。
依据漆酶的催化特性已经开发出漆酶电极。它具有测量范围宽、准确度高、稳定性好、操作简便等特点。漆酶和葡萄糖脱氢酶构建的双酶电极能够检测到纳摩级和次纳摩级的肾上腺素。20世纪90年代初,漆酶作为标记酶首次出现在免疫检测中。而漆酶更有望替代辣根过氧化物酶而成为新的标记酶。以漆酶为标记酶,其抗原-漆酶的接合灵敏度有很大提高,同时对培养基中变价金属离子的含量具有更低的敏感性。
7.6 饮料和食品中的应用
漆酶主要用于饮料澄清与色泽控制。它能氧化多酚类物质生成多酚氧化物,进而发生再聚合,形成可以被滤膜截留的大颗粒,达到净化饮料的目的。由于这一催化反应专一性强,污染少,澄清的果汁色浅且稳定,可以解决果汁加工中的二次混浊问题。另外,Pertersen利用漆酶中铜离子的催化特性,除去色拉、蛋黄酱等食品中溶解的氧,消除食品中的亚麻酸和溶解氧反应生成的挥发性异味组分,可达到改良食品感官参数的目的。
此外,漆酶还可用来测定食品中抗坏血酸的总含量。Huang发现邻苯二胺能被漆酶氧化成2,3-二氨基吩嗪,而L-抗坏血酸对2,3-二氨基吩嗪的生成具有抑制作用,漆酶酶活与抗坏血酸含量呈反比。这种方法快速、经济,而且能够测定连续上样的不同样品中抗坏血酸总量。该方法已应用在白酒、啤酒、奶粉等食品的检测中。
7.7 食用和药用菌生产
在食用和药用菌制种过程中,加入漆酶制剂能加速木质素的分解,为菌丝提供更丰富的养料,加快菌丝吃料,缩短培养时间。漆酶可作为食用菌和药用菌菌体呼吸作用中的末端氧化酶之一,有利于食用菌和药用菌呼吸过程中电子传递的正常运行,加大呼吸强度,为菌体的生命活动提供更多的可利用能量从而加速食用菌和药用菌的生长发育。漆酶分解木质素为菌体生长提供养料的同时,会产生酚类或醌类化合物,而酚或醌是一种很好的杀虫剂,能够抑制杂菌生长,防止食用菌和药用菌在制种和袋料栽培过程中的杂菌污染。漆酶在香菇袋料栽培过程中的转色阶段起着重要作用,影响香菇的产量和品质。利用这种方法将有益于节约木材和扩大生产规模。漆酶还有参与子实体形成的调控作用。
7.8 高分子化合物合成
漆酶能催化生漆中的漆酚氧化,产生高分子聚合物,使生漆干燥成膜,这是漆酶最重要的作用。
Murat Gure等研究表明,在水溶液中利用Trametes versicolor漆酶催化联苯三酚或其他酚类物质的聚合是获得具有新型结构聚合物的有效途径。由于取代苯酚类物质结构的特殊性,一般很难采用化学方法使其聚合,而用漆酶在缓冲溶液-有机溶剂的混合体系中催化对苯基苯酚的聚合却得到了高分子量的聚酚产物。Bruno等实现了在漆酶/O2的催化下,苯酚和含长链烷基的苯酚衍生物的有序聚合。Ikeda等在
研究表明,漆酶在真菌色素合成中起着重要的作用,并且可能参与菌索的形成,它也能在细胞间催化酚类化合物的聚合粘连来固定菌丝。人们已经利用漆酶的这种粘合功能进行化工产品的开发研究。漆酶可催化酚类、芳胺类和羧酸类等单体的聚合反应,在树脂材料、光电材料和染色工业等方面,都有着广阔的应用前景。
7.9 能源领域
生物燃料电池是利用酶或者微生物组织作为催化剂,将燃料的化学能转化为电能的。近年来,科学工作者致力于开发无隔膜的酶生物燃料电池。目前这类电池的工作寿命较短,所以还不适合于实际应用,仍需进行深入细致的研究。
7.10 生物肥料
近几十年来,加强木质纤维转化为腐殖质一直是国内外研究的重点。传统上应用高温堆肥的方法使农田秸秆转化为有机肥料,但是白腐菌产生的木质素降解酶系为堆肥产品的快速腐熟提供了实际的可能。目前,生物肥料的应用从培养参数的确定、突变品系的筛选及混合菌剂的联合利用等方面展开研究。
7.11 其他方面的应用
漆酶添加在漱口水、牙膏等各种口腔用品中可起到抑制口臭的作用,固定化漆酶可除去饮用水和灌溉用水中的有毒酚类化合物,净化生活和工业用水,减少环境污染。因为漆酶在氧化-还原过程中消耗氧气,添加于含油食品中可去除其中的氧气,防止食品氧化变质。漆酶可促使制糖甜菜中的果胶发生交联,生成食用凝胶。Lacki和Duvnjak利用漆酶处理低芥酸菜籽产品,3h酚含量降低约90%,使其可代替大豆蛋白用于食品和饲料的生产。漆酶还可催化染色剂中酚或芳胺类化合物在毛发上氧化成醌或黑色素使毛发染成黄色或黑色。
8 结束语
从漆酶发现至今已有100多年,经过各国科学工作者坚持不懈的努力,漆酶的研究取得很大的进展。随着研究的不断深入,人们对白腐真菌产酶机理、酶活性中心及作用机理都有了较多的了解,工业应用也已取得了较大进展,但仍存在不少问题,如菌株合成漆酶水平偏低,菌体生长较慢,酶催化效率低,不适于工业化生产等。我们现在应该考虑的是如何使漆酶的应用更加实用化,如何拓展漆酶新的应用领域和应用途径,将需要生物、化学、食品、农林等多学科的交叉交流,以及众多科研工作者们的不断努力。相信在不久的将来,漆酶在多个领域将会发挥更大的作用。
(参考文献略,需者可函索)
上一篇:酶制剂的酶活测定方法及影响因素
下一篇:β-半乳糖苷酶诱导条件优化的研究
分享到:
 收藏
收藏
收藏
评论排行
- ·中国发酵企业数据库(4)
- ·(4)
- ·CoQ10高产菌株选育的研究进展(2)
- ·抗生素发酵工艺所用冷却塔的性能分析及处理(1)
- ·微生物菌种选育技术.rar(1)
- ·发酵生产染菌及其防治(1)
- ·赤藓糖醇发酵工艺研究(1)
- ·重组AiiA 蛋白可溶性表达及发酵条件优化(1)
- ·生物反应器设计软件_发酵罐绿色版(1)
- ·酵母粉、酵母浸粉的区别(1)
- ·雷帕霉素研究进展(1)
- ·透明质酸用途和行业概况(1)
- ·黄酒制作工艺(1)
- ·水解(酸化)工艺与厌氧发酵的区别(1)
- ·糖蜜酒精废液处理过程中产生的微生物蛋...(1)
- ·紫杉醇高产菌发酵产物的分离、纯化和鉴定(1)
