生物柴油专用脂肪酶的开发及其应用研究
2007-12-18 21:45:31 来源:本站原创 评论:0 点击:
【关键字】 生物柴油专用脂肪酶;生物柴油;转化率
脂肪酶是一类特殊的酯键水解酶,脂肪酶作为生物催化剂可催化由不同底物出发的水解和合成反应。它能催化天然底物油脂(甘油三酯)水解,产生脂肪酸和甘油,在水解过程中存在中间产物甘油单酯和甘油三酯[1]。脂肪酶在催化合成生物柴油的反应中,具有举足轻重的作用,同化学法合成生物柴油相比,具有反应条件温和、副产品分离工艺较为简单,废水少,设备要求低等优点。然而不同来源的脂肪酶具有脂肪酸特异性,对甘油酯中Sn-1(3)和Sn-2位酯键具有不同的位置特异性,以及立体特异性[2]。如圆弧青霉脂肪酶对短链(C8以下)脂肪酸,黑曲霉和根霉对中等长链(C8~C12)脂肪酸表现出强烈特异性[1];Achromobaterium lipolyticum对饱和脂肪酸表现出特异性[3],而来源于Staphylococcus asreus226脂肪酶对不饱和脂肪酸具有特异性[4];白地霉对油酸甘油酯表现出强特异性,猪胰脂肪酶能够很好的催化甘油三酯中C18:1和C18:0同时对催化较多的碳链具有专一性[5,6]。因此在生物酶法合成生物柴油方面,研究出一种高转化率,对不同油脂原料具有广泛的适应性的生物柴油专用脂肪酶,具有非常重要的实际意义。
本文主要基于脂肪酶催化反应的特异性理论,根据不同油脂中的脂肪酸的种类,研究复配出生物柴油专用脂肪酶,催化不同的油脂具有高脂肪酸甲酯转化率即对不同原料具有广泛的适应性;根据不同油脂中的脂肪酸的种类不同,使用GC图谱对其产物进行分析复合酶催化4种油脂合成生物柴油(FAME)过程中脂肪酸甲酯转化率高于单酶催化的主要原因。
1 实验部分
1.1 实验主要材料
大豆油、棕榈油、菜籽油、花生油均购置广西南宁五里亭市场,化学试剂均天津美宇化学试剂有限公司,PB011脂肪酶、微生物脂肪酶、胰脂肪酶均由广西南宁庞博生物工程有限公司提供。
1.2 实验主要仪器
气相色谱仪(美国Aglilent科技公司),低速台式离心机(上海安亭科学仪器厂),微量移液器(上海求精玻璃仪器厂),恒温水欲振荡器(哈尔滨市电子技术开发公司)。
1.3 实验方法
HP-1非极性毛细管柱;FID检测器,H2流速:36.0ml/min,O2流速:360.0ml/min,N2流速:36.0ml/min,分流比1:50,恒定柱压:2.00psi,初始流速:1.7ml/min;程序升温:初始
气相色谱分析脂肪酸甲酯的含量,脂肪酸甲酯的转化率=样品中脂肪酸甲酯的含量/油脂完全甲酯化后脂肪酸甲酯的含量×100%。
称取经过适当处理的棕榈油,于50mL的锥形瓶中,反应为优化过的条件:称取棕榈油
在50mL三角瓶中加入一定量的不同底物、有机溶剂和复合脂肪酶,分三次流加甲醇,其中复合酶的配比为实验
2 结果与讨论
2.1 生物柴油复合酶的配比
表1 {3,3}单纯形格子点试验结果
|
试验号 |
X1 |
X2 |
X3 |
FAME |
试验号 |
X1 |
X2 |
X3 |
FAME |
|
1 |
1 |
0 |
0 |
52.00% |
6 |
2/3 |
0 |
1/3 |
46.95% |
|
2 |
0 |
1 |
0 |
38.37% |
7 |
1/3 |
0 |
2/3 |
28.00% |
|
3 |
0 |
0 |
1 |
0% |
8 |
0 |
2/3 |
1/3 |
25.80% |
|
4 |
2/3 |
1/3 |
0 |
43.97% |
9 |
0 |
1/3 |
2/3 |
12.61% |
|
5 |
1/3 |
2/3 |
0 |
45.92% |
10 |
1/3 |
1/3 |
1/3 |
22.27% |
对以上数据进行回归,得到的回归方程,利用Excel中的“规划求解”可以得到回归方程在X1=0.86,X2=0,X3=0.14(其中X1为PB011脂肪酶,X2为微生物脂肪酶,X3为胰脂肪酶)脂肪酸甲酯转化率最高。可见,各种脂肪酶的理论配比是,PB011脂肪酶占86%,微生物脂肪酶为0%,胰脂肪酶为14%。
2.2 复合酶和单酶催化大豆油的性能比较与分析
脂肪酸的甘油酯是植物油的主要成分,大豆油的主要是由棕榈酸C16:0、油酸C18:1(9)、亚油酸C18:2(9、12)、硬脂酸C18:0和亚麻酸C18:3(9、12、15)的甘油酯构成。图1复合酶催化大豆油的GC色谱图。
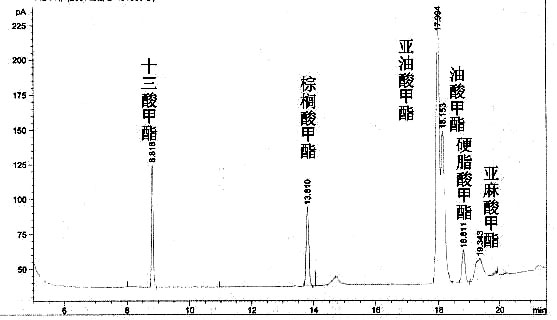
图1 复合酶催化大豆油的气相色谱图
从脂肪酶的催化的特异性理论我们可以知道,PB011脂肪酶是来源于白地霉,对油酸、亚油酸有很高的催化特性,微生物脂肪酶对sn-1(3)有很强的特异性;胰脂肪酶对cis-C18:1和cis-C18:0能够很好的催化,同时对催化较多的碳链具有特异性。从表2可以看出:PB011脂肪酶催化大豆由产物中油酸和亚油酸甲酯含量相对较高,硬脂酸甲酯含量偏低,亚麻酸甲酯几乎没有检测出;微生物脂肪酶的催化活性较低,对大豆油中各种脂肪酸的催化效率都很低;单一的胰脂肪酶催化大豆油,各种脂肪酸甲酯的含量比较低;PB011脂肪酶和胰脂肪酶酶构成的复合酶催化大豆油的GC图谱中,见图1。复合酶催化大豆油的GC图中比单酶催化大豆油的GC图谱中,多了一个峰,此峰代表的物质是亚麻酸甲酯,见图1。可见复合酶催化大豆油脂肪酸甲酯转化率的提高主要在于能够催化油脂中含有少量的亚麻酸,见表2。因此,此复合酶能够很好的催化以大豆油为原料油脂来合成生物柴油,比单酶催化FAME转化率明显提高了。
表2 3种单一脂肪酶和复合酶催化大豆油的脂肪酸甲酯转化率的比较
|
|
棕榈酸 |
亚油酸 |
油酸 |
硬脂酸 |
亚麻酸 |
总转化率 |
|
PB011脂肪酶 |
12.14% |
49.27% |
26.09% |
4.53% |
没有检测出 |
92.04% |
|
微生物脂肪酶 |
0.74% |
5.00% |
2.93% |
0.25% |
没有检测出 |
8.91% |
|
胰脂肪酶 |
6.38% |
23.62% |
12.91% |
2.57% |
没有检测出 |
45.48% |
|
复合酶 |
11.78% |
45.58% |
26.23% |
5.03% |
10.27% |
98.9% |
2.3 复合酶和单酶催化棕榈油性能比较与分析
复合酶和单酶分别催化棕榈油的结果见表3,可以看出PB011脂肪酶对棕榈油催化的效率最高,脂肪酸甲酯的转化率达92.33%;微生物脂肪酶对棕榈油几乎无活性,几乎没有脂肪酸甲酯检测出。当复合酶催化棕榈油,比单一的PB011脂肪酶催化棕榈油,其脂肪酸甲酯的转化率提高了将近3个百分点,可见,此复合酶对以原料为棕榈油的生物柴油的生产具有很好的适应性。图2是复合酶催化棕榈油的GC图谱,从图谱中,可以看出,复合酶催化棕榈油脂肪酸甲酯转化率的提高主要在于,能够催化棕榈油中含有的亚麻酸。
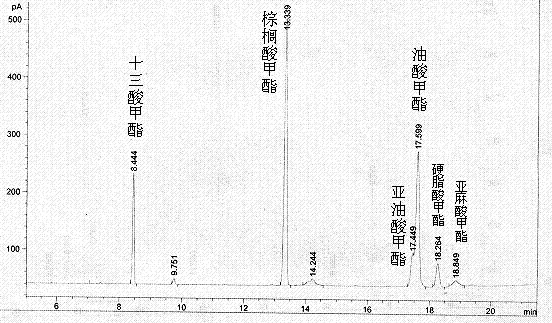
图2 复合酶催化棕榈油的气相色谱图
表3 3种单一脂肪酶和复合酶催化棕榈油的脂肪酸甲酯转化率的比较
|
|
棕榈酸 |
亚油酸 |
油酸 |
硬脂酸 |
亚麻酸 |
总转化率 |
|
PB011脂肪酶 |
42.20% |
7.91% |
37.37% |
4.85% |
没有检测出 |
92.33% |
|
微生物脂肪酶 |
0 |
0 |
0 |
0 |
没有检测出 |
0 |
|
胰脂肪酶 |
28.12% |
5.19% |
21.58% |
2.26% |
没有检测出 |
57.15% |
|
复合酶 |
48.16% |
6.09% |
33.04% |
5.59% |
2.35% |
95.23% |
2.4 复合酶和单酶催化花生油的性能比较与分析
表4是复合酶和单酶催化花生油的实验结果,可以看出,单一的PB011脂肪酶和微生物脂肪酶催化花生油合成生物柴油的过程中,没有亚麻酸甲酯的生成,胰脂肪酶催化花生油得到的生物柴油产品中虽然有亚麻酸甲酯的生成,但是含量相对较低,而此复合酶催化花生油,亚麻酸甲酯含量相对较高。3种单一脂肪酶和复合酶催化花生油的脂肪酸甲酯转化率的定量情况见表4。
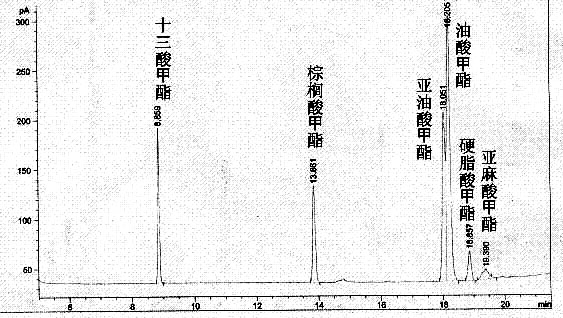
图3 复合酶催化花生油的气相色谱图
表4 3种单一脂肪酶和复合酶催化花生油的脂肪酸甲酯转化率的比较
|
|
棕榈酸 |
亚油酸 |
油酸 |
硬脂酸 |
亚麻酸 |
总转化率 |
|
PB011脂肪酶 |
12.34% |
30.12% |
43.36% |
4.27% |
没有检测出 |
90.12% |
|
微生物脂肪酶 |
4.80% |
18.17% |
32.82% |
1.51% |
没有检测出 |
57.30% |
|
胰脂肪酶 |
7.69% |
19.31% |
24.94% |
2.58% |
没有检测出 |
54.52% |
|
复合酶 |
13.22% |
28.51% |
47.73% |
4.88% |
3.45% |
97.8% |
从表4可以看出,复合酶催化花生油合成生物柴油的过程中,脂肪酸甲酯转化率的提高主要是由于复合酶中的胰脂肪酶能够催化花生油中的亚麻酸从而条了脂肪酸甲酯的转化率,见图3。复合酶催化花生油,其中脂肪酸甲酯的转化率可达97.8%。可见,复合酶对花生油具有较好的催化性能。
2.5 复合酶和单酶催化菜子油性能比较与分析
目前国内对酶法催化菜籽油合成生物柴油过程中,脂肪酸甲酯的转化率普遍比较低,基本在80%左右。主要是因为在菜子油中含有较多的20碳稀酸和芥酸而脂肪酶对C18以上的碳链很难催化,以至影响脂肪酸甲酯的产率。
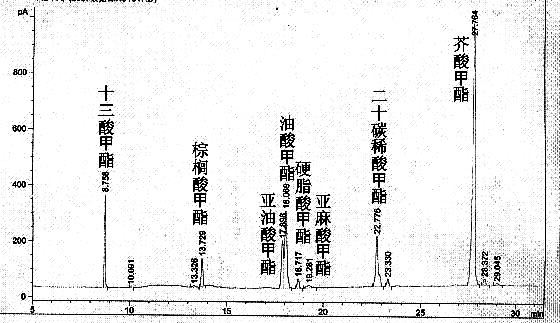
图4 复合酶催化菜子油的气相色谱
从表5看出,微生物脂肪酶对菜子油几乎无催化特性,PB011脂肪酶对菜籽油虽然有催化特性,但是无法催化菜子油中的20碳稀酸和芥酸,故其脂肪酸甲酯转化率仍然很低。胰脂肪酶单独催化菜子油的时候,并不能说明其特异性,在胰脂肪酶和PB011脂肪酶构成的复合酶催化菜子油的GC色谱图中,见图4,可以看出,多了3个峰,分别代表的产物是亚麻酸甲酯、20碳稀酸甲酯和芥酸甲酯,可见利用复合酶催化菜子油合成生物柴油时在很大程度上,催化了菜子油中的20碳稀酸和芥酸,主要是由于复合酶中的胰脂肪酶对cis-C18:1和cis-C18:0具有很好的催化性能,同时对催化较多的碳链具有特异性,从而提高了脂肪酸甲酯的转化率。表5列出了3种单一脂肪酶和复合酶催化菜子油的脂肪酸甲酯转化率情况。
表5 3种单一脂肪酶和复合酶催化菜子油的脂肪酸甲酯转化率的比较
|
|
棕榈酸 |
亚油酸 |
油酸 |
硬脂酸 |
亚麻酸 |
20碳稀酸 |
芥酸 |
总转化率 |
|
PB011脂肪酶 |
2.84% |
8.24% |
13.42% |
1.07% |
0 |
0 |
0 |
6, 5.57% |
|
微生物脂肪酶 |
0.51% |
1.33% |
2.10% |
0.17% |
0 |
0 |
0 |
4.1% |
|
胰脂肪酶 |
0.81% |
5.45% |
9.45% |
1.12% |
0 |
0 |
0 |
48.33% |
|
复合酶 |
3.02% |
9.48% |
15.65% |
1.25% |
0.92% |
8.72% |
49.03% |
88.09% |
从表5可以看出,在单酶催化菜籽油时,各种脂肪酶的单一催化作用,表现出极低的催化性能,脂肪酸甲酯的转化率非常低,最高只达65%左右。在此复合酶催化菜籽油时发挥了单酶无法具有的催化特性即从一定程度上消除了对底物的特异性。最后测得脂肪酸甲酯的转化率高达88.09%比单酶催化,脂肪酸甲酯的转化率提高了20个百分点。
3 结论
(1)根据不同来源的脂肪酶对底物油脂的具有不同的催化特异性(脂肪酸特异性、立体特异性、双键位置特异性)着手,对复合酶的复配进行研究,以催化油脂合成生物柴油过程中的脂肪酸甲酯转化率为指标,最后确定复合酶的的配比为:PB011脂肪酶为86%,胰脂肪酶为14%。
(2)通过研究得到的生物柴油专用脂肪酶在优化的条件下,分别催化大豆油、棕榈油、花生油、菜籽油,最后测得的FAME的转化率分别高达:98.9%、95.23%、97.8%、88.09%。
(3)通过3种单一的脂肪酶和复合酶催化4种常用的油脂,以脂肪酸甲酯的转化率为指标的对其性能进行了对比,从GC分析结果,解释了复合酶在催化油脂过程中,表现出单一酶所不具备的催化特性原理,突破了一种脂肪酶催化不同的油脂原料具有较高的转化率,开发出了一种,对不同的原料(油脂)的都具有高转化率生物柴油专用脂肪酶,弥补了当前国内外对酶法合成生物柴油的中“生物柴油专用脂肪酶”的空缺。
参考文献
[1] 李香春,甄宗园.脂肪酶特性及其应用[J].粮食与油脂,2003,3:19-20.
[2] 李燕,潘运国,连毅.微生物脂肪酶催化及其性质研究进展[J].2005,10:15-17.
[3] Davranov K.Microbial lipases in biotechnology[J].Appl.Biochem.Microbiol.,1994,30:
527-534.
[4] Muraoka T, ando T, Okuda H. Purification and properties of a novel lipase from Staphylococus aureus 226[J].Biochem.,1982,92:1933-1939.
[5] 曹淑桂.脂肪酶的底物特异性及其应用潜力[J].生物化学与生物物理进展,1995,22(1):9-11.
[6] 邬显章,邬敏辰.脂肪酶分子生物学的研究进展[J].食品与生物技术,2002,21(1):94~95
[7] 庾乐.有机相酶催化制备生物柴油的研究[D].2006,广西大学硕士研究生学位论文,18-19.
[8] 李云雁,胡传荣.试验设计与数据处理[M].北京:化学工业出版社,2005.
刘汉灵,男,44岁,广西南宁人,博士,副教授,广西大学硕士生导师,广西南宁庞博生物工程有限公司总经理,主要从事新型酶制剂的研究与开发。
相关热词搜索:α-乙酰乳酸脱羧酶
上一篇:木聚糖酶产生菌的选育
下一篇:黄孢原毛平革菌产木素过氧化酶的研究
分享到:
 收藏
收藏
收藏
评论排行
- ·中国发酵企业数据库(4)
- ·(4)
- ·CoQ10高产菌株选育的研究进展(2)
- ·抗生素发酵工艺所用冷却塔的性能分析及处理(1)
- ·微生物菌种选育技术.rar(1)
- ·发酵生产染菌及其防治(1)
- ·赤藓糖醇发酵工艺研究(1)
- ·重组AiiA 蛋白可溶性表达及发酵条件优化(1)
- ·生物反应器设计软件_发酵罐绿色版(1)
- ·酵母粉、酵母浸粉的区别(1)
- ·雷帕霉素研究进展(1)
- ·透明质酸用途和行业概况(1)
- ·黄酒制作工艺(1)
- ·水解(酸化)工艺与厌氧发酵的区别(1)
- ·糖蜜酒精废液处理过程中产生的微生物蛋...(1)
- ·紫杉醇高产菌发酵产物的分离、纯化和鉴定(1)
