微生物发酵生产花生四烯酸的研究进展
2007-05-13 14:02:19 来源:中 国 油 脂 评论:0 点击:
1 花生四烯酸概述
1 1 花生四烯酸的结构和理化特性花生四烯酸(ArachidonicAcid,AA)系统命名为全顺-5,8,11,14-二十碳四烯酸,又名5,8,11,14-花生酸,分子量304 5,分子式C20H32O2,结构式如图1所示。
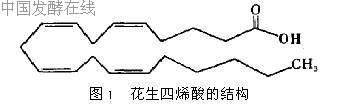
花生四烯酸在室温下呈液体,其熔点-49 5℃,沸点245℃(分解),溶解于醇、醚和水中,碘值333 50gI/100g,紫外吸收峰257、268、315nm[1]。
1 2 花生四烯酸的代谢途径和生理功能AA是ω-6系列多不饱和脂肪酸,主要以磷脂的形式存在于机体各种组织的细胞膜上,浓度通常小于10-6mol/L,决定着细胞膜的一些重要生物活性。细胞膜磷脂可在磷脂酶A2和磷脂酶C的作用下释放出AA。在哺乳动物中,AA通常是亚油酸代谢得到,途径为:食物来源的亚油酸先脱饱和生成γ-亚麻酸(GLA),再经延长碳链,脱饱和生成AA。然后AA再转变成前列腺素,白三烯,血栓素等类二十烷[2]。AA和这些代谢产物具有很强的生物活性,如参与神经内分泌,调节平滑肌收缩,促进细胞分裂,抑制血小板聚集等等。近年来研究发现,AA具有保护皮肤,提高免疫能力,促进胎儿发育,促进生物体内脂肪代谢,降低血脂、血糖、胆固醇等功能,已引起人们的高度重视[3],AA是一种人体必需的多不饱和脂肪酸。通常情况下,人体内的AA可由食物中的亚油酸来合成,但对于老年人、婴幼儿以及高血脂症、糖尿病等患者来说,其体内Δ6,Δ5-脂肪酸脱饱和酶的活性较弱或受到抑制,从而造成AA缺乏,导致机体代谢紊乱而引发多种疾病[4]。
1 3 花生四烯酸的应用正因为AA在降血脂、抑制血小板聚集、抗炎症、抗癌、抗脂质氧化、促进脑组织发育等方面具有独特的生物活性,它已在食品、医药、化妆品等领域得到广泛应用。目前,AA通常是和其他多不饱和脂肪酸一起制成软胶囊用于预防和治疗湿疹、皮炎和高血压,脑血栓、冠心病、动脉硬化等心脑血管疾病。另外,以AA或亚麻酸为原料得到前列腺素,是抗血栓、扩血管新药,广泛用于治疗血栓性脉管炎,视网膜中央静脉血栓,保护急性心肌缺血等[1]。
在功能性食品方面,AA和其他多不饱和脂肪酸被誉为“21世纪功能性食品的主角”,可作为食品添加剂或营养强化剂,改善人类的膳食结构,增强机体的免疫力。添加了PUFA的婴儿奶粉、饮料、果冻、饼干、糖果在国外已经十分普遍。日本资生堂将PUFA加到化妆品中,制成护肤护发防皱等美容用品,起到保护皮肤水分,延缓皮肤老化,治疗慢性湿疹的功效。据FDA1992年报道,有29种化妆品中含有AA成分。新近研究发现,人和动物中的酶系统在将饮食中α-亚麻酸及亚油酸转化成AA、GLA、DHA、EPA的能力是有限的,如果在动物饲料中添加含PUFA产生菌的菌体干粉,既可提高饲养效率,又能增加肉蛋奶中PUFA的含量[1]。
2 花生四烯酸发酵的研究
2 1 花生四烯酸的来源动物的肾上腺、肝脏、沙丁鱼油,蛋黄中含有AA,但含量很低,一般小于0 2%(W/W)[1]。因此,国外很早就开展了微生物生产AA的研究,现在日本的SuntoryLtd ,美国的MartekBiosciencesCo ,瑞典的Hoffmann-LaRocheLtd 等[5]公司已经实现大规模工业化生产。目前,国内有数家单位也已经开始发酵试生产。1999年华南理工大学[4]对被孢霉Mortierellasp M10进行了5L罐发酵放大实验。2000年中科院等离子体物理研究所[6]报道了利用高山被孢霉(Motiellaalpina)30、250L发酵罐实验结果。2002年武汉烯王生物工程有限公司[7]报道了利用(M alpina)进行50t罐工业化生产的研究。
2 2 高产菌株的筛选和改良
2 2 1 产花生四烯酸的微生物 某些原生动物如阿米巴,显微藻类如红藻、蓝藻,部分细菌、酵母菌和多数丝状真菌均可以产生AA,但其产生量足以希望用来作为发酵菌种的却不多,从国内外文献上来看,绝大多数研究集中在藻状菌纲:Conidiobolus(耳霉属),Mortieella(被孢霉属),Mucor(毛霉属),Rhizopus(根霉属),Diasporangium(枝霉属)等。尤其以高山被孢霉(M alpina)、长被孢霉(M elongata)、深黄被孢霉(M isabellina)、拉曼被孢霉(M ramanni ana)、终极腐霉(Pythiumultimum)为最。
2 2 2 筛选和改良 国外这部分工作是在20世纪80年代完成的。1987年山田等[8]人从土壤中分离到多株AA产生菌,经选育获得1株高产菌高山被孢霉(M alpinaIS-4),它用葡萄糖作碳源时产生4 3g/LAA。1997年曲阜师范大学杨革等[9]人从14株丝状真菌出发筛选出U-7,然后经过紫外线(UV)氯化锂(LiCl)复合诱变得到U-71,其PUFA总含量为27 5%,AA含量为0 7%。1998年福建师范大学黄建忠等[10]人以深黄被孢霉(M isabellinaAS3 3410)为出发菌株,经UV、DES(硫酸二乙酯)、NTG(亚硝基胍)复合诱变选育出M018,摇瓶发酵油脂含量比出发株提高133%。杨革等[11]人从土壤中分离筛选出1株拉曼被孢霉(M ramannianaSM541),并以此为出发菌株经过UV、LiCl复合诱变得到SM541-9,其AA的含量从321mg/L提高到623mg/L。1999年华南理工大学朱法科等[12]人以1株产AA的被孢霉为出发菌株经UV诱变选育了突变株Mortierellasp M10,其干菌体得率为31g/L,油脂含量为8 3g/L,气相色谱分析AA占总脂的10 06%。2000年华中理工大学周蓬蓬等[13]人利用DES对高山被孢霉M3-18的原生质体进行处理,得到AA产量比对照组提高4 4倍的突变型M20。中科院等离子体物理研究所[6]对M alpina采用离子注入法诱变得到1株AA高产菌,其每升培养液可得生物量30 80g,干菌体油脂含量为25 8%,AA的含量占总脂的45 37%。2001年武汉工业学院何东平等[14]人对11株产脂丝状真菌通过拟定的7项指标筛选诱变出一产脂优良株,并进行了中试发酵研究,AA含量达4 6%。
在分子水平上的研究,主要集中在对脂肪酸脱饱和酶的序列分析和基因的遗传操作上,以控制生物体膜脂与贮脂中的PUFA的含量。理论上可以通过增加限速酶基因拷贝数或定点突变技术来消除反馈调节和代谢支路调节,提高基因表达能力;还可以通过引入克隆基因的反义表达载体来封闭基因表达,增加中间脂肪酸的产量[15];也可尝试采用新兴的人工分子进化技术———DNAshuffling技术来快速获得相关基因的正突变。国外在这方面鲜有报道,国内主要是华南理工大学和南开大学开展了部分工作。2000年华南理工大学张羽航等[16]人构建了被孢霉cDNA文库,并以此筛选出Δ9-脱饱和酶cDNA序列。南开大学李明春等[17]人利用CHEF凝胶电泳技术分析了深黄被孢霉(M isabellinaAS3 3410)、拉曼被孢霉(M ramannianaAS3 3410)、葡酒色被孢霉(M vinaceaAS3 3410)的电泳核型。2001年南开大学刘莉等[18]人从含有高山被孢霉(M alpinaATCC16266)Δ6-脱饱和酶基因的重组质粒中酶切出目的片断,通过大肠杆菌和酿酒酵母的穿梭载体,最后得到酿酒酵母工程种YMAD6,其目的基因表达量超过了国际最高水平。
代谢调控育种方面,可以考虑从以下几点出发:切断或减弱支路代谢,如选育α-亚麻酸、EPA、DHA缺陷型或渗漏型,解除自身反馈调节。据报道,选育4-氟代亚油酸抗性突变株可使被孢霉的AA产量提高0 65倍,还可以选育耐高浓度AA的突变株,增加前体物的合成。报道称,选育脱氢赖氨酸敏感、萘啶酮酸敏感、氟乙酸敏感的突变株均可增加AA的产量;另外,选育Δ5-脱饱和酶活性强的突变型、低温生长突变型、优良产脂突变型,强化能量代谢等均有利于AA产量的提高[19]。
2 3 培养条件的研究目前,国外的研究多集中在这一部分。1991年PramodKBajpai等[20]人研究了M alpinaATCC32222在25℃培养时葡萄糖浓度和起始pH对AA产量的影响,发现当pH在6 0~6 7,葡萄糖浓度在100g/L时有利于AA的积累。1997年SinghA等[21]人发现用玉米浆代替硝酸钠,添加豆粉和植物油,均能提高M alpinaATCC32222产AA的能力。1998年日本HigashiyamaK等[22]人采用M alpinaIS-4,添加适量的KH2PO4、Na2SO4、CaCl2和MgCl2,1万L罐发酵8d,AA的产量达10 8g/L;接着又研究了溶解氧对该菌种形态和AA产量的影响,并得出了最佳值[23]。2000年SredanskyM等[24]研究P ul timum的固体培养,每克湿培养基产AA2 6mg。1997年华南理工大学鲍时翔等[25]人采用正交试验对Morrierellasp M10进行培养基和发酵条件的优化,其发酵结果是,细胞干重和AA产量分别为33 51g/L和0 827g/L。接下来又对该发酵过程进行分析,建立了菌体生长、油脂形成的动力学模型,应用实验数据回归了模型参数,预测值与实验值有较好的吻合[26]。1999年鲍时翔等[4]人又研究了碳源种类、葡萄糖浓度、添加植物油对AA产量的影响,并进行了5L罐放大试验,细胞干重和AA产量分别提高到38 4g/L和0 95g/L。中国热带农业科学院黄惠琴等[27]人考察了温度、葡萄糖加料方式、菌体老化对AA产量的影响,结果是变温培养,分批加料,菌体老化有利于AA的积累。2000年南京师范大学刘吉华[28]对1株产AA的轮枝霉(Diaspo rangiumsp )进行了碳源氮源种类、pH、温度、接种量、接种龄、通气量等培养条件的优化,结果AA产量达549 8mg/L。2002年华中科技大学余龙江等[29]人研究了不同浓度谷氨酸对高山被孢霉M3-18菌株的影响,发现发酵第7d时总脂和AA产量达最高值,生物量、油脂和AA产量别为24 43、9 21、1 41g/L,分别比对照组提高了1 13、1 15和1 69倍。2002年武汉烯王生物工程公司尚耘等[30]人研究了温度、pH、培养时间等对高山被孢霉的细胞生长、油脂合成和AA积累的影响,确定了最佳培养基和条件,使得其干菌体重达到25 9g/L,总脂含量为36 6%,AA为71 0%。
2 4 花生四烯酸的提取

AA是胞内产物,因其产量的高低受到菌体得率、总脂回收、AA得率等因素的制约,而且AA在油脂中的绝对含量低,毛油成分复杂,在分离纯化过程中极易受到氧、光、热等的作用而发生氧化、聚合、降解、异构等反应,产生对人体有害的成分,因此分离提取工艺显得十分重要,尤其是油脂的精炼和AA的提取这两个环节。2000年何东平等[31]人建立了水化脱胶、碱炼脱酸、脱色、脱臭4步精炼工艺,所得的微生物油脂经过色泽、气味、杂质、酸值、加热试验、含皂量等多项指标的检测达到食用标准。AA的富集可以采用低温结晶、尿素包合、分子蒸馏、吸附分离、超临界流体萃取、脂肪酶浓缩等方法[32]。2002年袁成凌等[33]人以甲醇为溶剂,脂肪酸为脲包客体,在-10℃的条件下用尿素包合法富集微生物油脂中的AA,经气相色谱分析其浓度从38 29%提高到78 97%,总收率达到90%以上。
3 问题和展望目前,亟待解决的问题是,继续筛选高产菌株和改良现有菌种以提高AA的产量,并进一步优化发酵条件,建立高效的目标产物分离提取手段。在基础理论方面,要大力研究脂肪酸脱饱和酶及其基因的作用和调节机制,争取在分子水平上获得更加深刻的认识,以期采用遗传操作方法来改造菌株。
如今,心脑血管疾病已经成为威胁健康的严重杀手之一,而且人们的保健美容意识正变得越来越强,AA作为功能性油脂的一种重要成分,在食品、药品和化妆品行业的需求也必将大幅增加。我们有理由相信,开发成熟的AA生产过程,其社会经济意义极为重大。
相关热词搜索:花生四烯酸
上一篇:轮梗霉高产花生四烯酸的诱变育种研究
下一篇:真菌发酵法产油脂的研究
分享到:
 收藏
收藏
收藏
评论排行
- ·中国发酵企业数据库(4)
- ·(4)
- ·CoQ10高产菌株选育的研究进展(2)
- ·抗生素发酵工艺所用冷却塔的性能分析及处理(1)
- ·微生物菌种选育技术.rar(1)
- ·发酵生产染菌及其防治(1)
- ·赤藓糖醇发酵工艺研究(1)
- ·重组AiiA 蛋白可溶性表达及发酵条件优化(1)
- ·生物反应器设计软件_发酵罐绿色版(1)
- ·酵母粉、酵母浸粉的区别(1)
- ·雷帕霉素研究进展(1)
- ·透明质酸用途和行业概况(1)
- ·黄酒制作工艺(1)
- ·水解(酸化)工艺与厌氧发酵的区别(1)
- ·糖蜜酒精废液处理过程中产生的微生物蛋...(1)
- ·紫杉醇高产菌发酵产物的分离、纯化和鉴定(1)
