饲料液态酶及其后喷涂工艺的研究与应用
2010-06-28 15:52:15 来源:本站原创 评论:0 点击:
饲料安全是关系到食品安全和消费者利益的一件大事,是全社会关注的热点。实现畜产品“绿色化”的核心问题是精确使用抗生素等药物类添加剂,更多地应用诸如酶制剂、微生物制剂等“绿色”生物技术产品,以解决畜牧生产中疾病的预防、生产性能的提高等问题。饲用酶制剂作为一种新型“绿色”环保添加剂,自从1975年美国饲料工业首次把酶制剂作为添加剂应用于配合饲料中并取得显著效果后,饲用酶制剂日益受到世界养殖业的重视。其效能特点有:
① 补充动物内源酶的不足,提高饲料报酬;
② 降解植物细胞壁,促进营养物质的消化吸收;
③ 消除饲料中的抗营养因子,提高饲料安全性;
④ 增强动物的抗病能力,提高畜禽成活率;
⑤ 降低氮、磷的排泄量,减少环境污染。
⑥ 降低饲料成本。
饲料酶按功能主要分为两类:一类是降解单底物的植酸酶、淀粉酶、蛋白酶、木聚糖酶、纤维素酶、葡聚糖酶、甘露聚糖酶等单酶,其中以植酸酶和木聚糖酶、甘露聚糖酶应用最普遍,多采用基因工程菌液体发酵生产。另一类是以非淀粉多糖为底物,真菌固体或液体发酵,进一步复配单酶加强的复合酶。
为了提高饲料酶的使用效果,需要从酶学性质改进和使用方法优化两方面研究。酶学性质包括酶活、底物适应性、pH曲线、温度曲线、结构稳定性等酶蛋白本身特征,主要由生产菌种和生产工艺决定。使用方法包括酶的剂型、储存稳定性、加工稳定性、酶的配比和剂量与作用饲料底物的对应降解效果。康地恩生物集团等酶制剂生产商建设现代化生产基地,构建高效工程菌株,改进发酵工程技术,配合颗粒饲料大力推广先进的液态酶后喷涂工艺,成为2008年饲料酶一大亮点。
一、基因工程菌株的构建
人们为了解决单胃动物不能利用植物中植酸磷、非淀粉多糖的问题而寻找并发现了植酸酶、木聚糖酶、纤维素酶等饲料酶。对这种应用性很强的酶所进行的基因工程研究紧紧围绕着工业化应用这一中心目标。基因工程饲料酶在养殖业中的应用,有三种可能的途径。
第一,通过植物基因工程改造饲料用作物,直接在植物的籽实中表达合适的植酸酶,省去了植酸酶的生产及其在饲料中的添加。Pen等(1993)首先在烟草种子中成功地表达了外源基因植酸酶,植酸酶的表达量达到种子中可溶蛋白的1%、叶片可溶性蛋白的14.4%[1]。Li J于1997年在大豆(Glycine max)的籽实中成功表达来自黑曲霉的植酸酶,并且其酶学性质与出发菌株所产的相同;饲喂实验结果表明同样可以提升家禽对植酸磷的利用率[2]。近年来,在水稻、苜蓿、玉米、油菜中成功表达植酸酶。但是由于植物籽实作为配合饲料的原料经过饲料生产的全过程,因此其中所包含的植酸酶必须经历高温制粒的过程。同时由于各地用作动物饲料的作物种类甚或品种往往不同,很难仅仅为了提高家畜家禽的植酸磷利用率而大规模弃用原来的优势品种。植酸是植物体内的第二信使之一,在植物生长发育过程中起着重要的作用,也是种子储存磷的主要机制,如果植酸酶过量表达,会对植物造成伤害,出芽率较低[3-6]。转基因植物可以表达具有活性的β-葡聚糖酶和木聚糖酶,并正常生长发育。作为结构物质的非淀粉多糖,如果非淀粉多糖相关酶的合成量提高,也会打破该植物整体的生理平衡,出现倒伏、减产[7]。
第二,利用现代转基因动物技术,使饲料酶基因在单胃动物消化道内源表达,无须外部添加。Serguei P等于2001年将ECP编码基因appA转化入小鼠中。55kDa的活性植酸酶蛋白通过唾液腺分泌入唾液中。转植酸酶基因小鼠的粪便中磷的含量与阴性对照相比降低了11%。Golovan SP等于2001年将植酸酶基因转入猪内,分泌含有植酸酶的唾液以分解食物中的植酸磷,它排出的粪便中含磷量与对照相比降低了75%。这方面的研究起步较晚,仍有许多问题有待解决,其中最主要的是转基因过程对动物体的损伤,及如何避免等问题[3-5]。
第三,微生物发酵生产饲料酶,作为固体饲料添加剂预混在粉状配合饲料中,或制备液体酶在高温制粒后喷涂于颗粒饲料表面。这种策略对使用者来说最为简单易行,但对植酸酶的生产者提出了很高的要求。在微生物体内,植酸酶并非持家基因,因此天然菌株生产植酸酶的能力十分低下,远远不能满足饲料成本控制的需要。野生型黑曲霉NRRL3135菌株产植酸酶的最高水平为6.8 U/ml,采用现代基因工程技术大幅提高生产植酸酶的能力是最为直接的方法。
第一个被分离纯化的植酸酶来源于Aspergillus terreus No. 9 A - 1,从此以后,陆续从十几种微生物中分离到植酸酶、木聚糖酶、甘露聚糖酶,如枯草芽孢杆菌、假单孢杆菌、乳酸杆菌、大肠杆菌、酵母、曲霉、隔孢伏革菌等,并对酶的生理生化性质进行了较深入的研究[8-11]。
国内以中国农科院饲料所姚斌等为代表的研究者构建了上述饲料酶的大肠杆菌、毕赤酵母等高效表达系统,并迅速产业化[12]。利用转基因技术构建的毕赤酵母表达系统与液体深层发酵工艺相结合,大大拓展了饲料酶基因的来源,为提高饲料酶的产量和酶活提供了强大的技术支持。国家高技术研究发展计划(863计划) “饲料用酶的分子改良与产品研制”立足饲料行业对酶制剂的急需,组织中国农科院、吉林大学、华东理工大学、康地恩、挑战等科研单位与企业,在酶分子改良、高效表达、工程菌高密度发酵、饲料酶规模化生产与应用技术等方面联合攻关,通过分子改良,将满足饲料酶高比活、耐酸性、抗胃肠道蛋白酶等要求,预期木聚糖酶、葡聚糖酶、甘露聚糖酶、半乳糖苷酶在工程菌中表达量高于5g/L,发酵液效价分别达到100000IU/mL、50000IU/mL、8000IU/mL、5000IU/mL。
植酸酶已成为21 世纪世界公认的、最有前景的无公害饲用酶制剂。1999 年植酸酶全球销售额为全部饲用酶制剂的49%。在德国和荷兰,超过90%的猪鸡饲料添加了植酸酶。欧美等国的植酸酶每年以20%左右的速度在递增,亚洲地区以每年80%~100%的速度在递增。中国是全世界首屈一指的饲料需求大国,2010年和2020 年,中国的饲料需求量分别为3.4 亿t 和4.08 亿t,植酸酶添加量以0.1‰计算,植酸酶纯蛋白的需求量应达到3.4~4.08 万t 以上,由此产生的直接经济效益每年将超过30亿人民币,具有良好的市场前景。
二、液态酶的生产与后喷涂工艺
酶作为一种蛋白质,对环境因素很敏感。温度、pH值、水分、强酸、强碱、紫外线和贮存过程等都会对酶活产生很大的影响。如上所述,分子改良的饲料酶基本适合饲料和消化道的作用环境,但随着大型饲料厂中越来越多地装备有强调质设备以及高温(80-150℃)短时加工设备。在饲料制粒、挤压和膨化过程中受温度、压力和水分的强烈作用下,酶制剂等热敏性微量组分的大部分活性都丧失了。康地恩检测深圳某公司提供的11个添加植酸酶的预混料样品,结果60天酶活平均降低50%。
将酶制剂进行“包被”处理,制成微胶囊,减少了外界因素的影响,酶的热稳定性也有所提高,但液态酶后喷涂工艺由于高均匀度及避免高温蒸汽对酶活的不利影响,越来越普及,成为颗粒饲料生产必备的配套设备。KDN液态植酸酶、木聚糖酶、复合酶和后喷涂设备相配套,在六和、新希望、中惠、双胞胎、正邦、海新、粤星、永达、得利斯等国内主要大型饲料企业推广应用,能显著降低饲料成本,提高消化率,减少氮磷排放,受到饲料企业的欢迎,已经安装300余台,对饲料酶的普及应用具有重要的推动作用。
1、理想的后喷涂设备要求
① 微量添加,高均匀度。用高压空气作载体,气液双料喷嘴,将微量(50-200ml/吨饲料)酶液充分雾化,弥漫包涂饲料颗粒;喷酶的颗粒饲料成品硬度无影响,水分几乎不增加;并辅设空气清洗系统,杜绝喷嘴处酶液滴漏问题。
② 自动化,高精确度。通过压力传感器连续测定干料流量,经PLC信息处理,自动控制酶蠕动泵流量,使酶的喷涂量准确跟踪干料流量变化。
③ 兼容性好。多液仓,多泵,多喷嘴,适合多酶种以及其他液体添加剂喷涂。各液仓设置传感器,方便监控酶液用量和盘点。
④ 触摸屏人机界面,总览全部过程,操作方便,参数设置简单,可根据饲料不同粒度选择相应的校正曲线。
⑤ 加缓存仓,使颗粒干料流量稳定,均匀度提高。
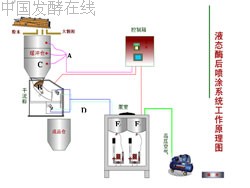
2、KDN-PPA液态酶后喷涂设备组成和工作原理
KDN-PPA按照上述要求,采用最先进自动化控制技术研制而成,主要的电控件、泵、传感器、阀均采用国际名牌产品,确保产品运行可靠,计量准确。KDN-PPA设备的硬件由泵室、添加室、控制箱组成:
① 泵室含有液仓、液体流量测定装置、液泵等。
② 添加室含有颗粒饲料流量传感器、喷涂装置等。
③ 控制箱含有PLC、触摸屏等。
合格的饲料颗粒经分级筛进入缓冲仓(C),控制箱内的PLC收到仓料料位传感器(A)的有料输入信号,产生输出,给控制箱内颗粒饲料流量表启动信号,根据人为指定的目标流量(t/h)进行PID调节,相应打开添加室进料门(B),饲料靠自重流下,同时,安装在添加室内的喷涂电磁阀(D)打开,泵室根据触摸屏中指定的酶制剂流量(1/h)、酶泵的选择以及颗粒饲料流量表的开门信号启动,将高压空气和酶(E和F)按要求的流量和压力送到添加室,喷涂到颗粒料。
3、高均匀度、高喷涂率
饲料主要以固体和液体形态存在,各种原料混合方式无非三种方式:固-固、固-液、液-液。
添加剂在颗粒饲料中分布均匀度受添加量、添加剂的粒度、颗粒饲料粒度、混合机性能等因素影响,设某原料的添加量为x(g/kg),添加剂颗粒度为y(粒/g),颗粒饲料粒度为z(粒/g),则每颗饲料含有原料的粒数为:
如固体植酸酶x=0.1, y=1500, z=15,则
2-3kg的高压空气,可以将0.1ml液体酶可以分散为0.8-6.4×106个雾滴。液体植酸酶x=0.1, y=3000000, z=15,则
对已经安装的后喷涂设备进行了30多台次的均匀度检测,平均变异系数CV=13%,喷涂效率(喷涂颗粒实际酶活占理论值的比率)85%。而植酸酶颗粒剂直接混合到颗粒饲料中,则混合过程酶活从418IU/mL降到217IU/mL,73.5°C调质后降到114 IU/mL,制粒后降到97IU/mL97,植酸酶的活性降低了75%,均匀度CV=39%(滨州六和实地测定数据)。
4、高保存率
在潍坊六和测定结果表明,植酸酶粉剂( 5897u/g)4倍添加于饲料(400g/t),经高温制粒(75°C),残留酶活很低,仅50U/kg,与理论值 2300u/kg和预期值 500u/kg差距甚远。与后喷涂效果450u/kg差距也很大。
由于液体酶喷涂于颗粒饲料表面,担心易脱落或不稳定。临沂六和模拟养殖场室外饲料筒仓处理:相对湿度75%,60℃保持10小时;37℃保持14小时交替处理,共处理10天,比较进口固体耐制粒植酸酶直接混合、KDN液体植酸酶后喷涂生产的颗粒饲料,分别存留41%和56%。
5、液体酶生产
KDN液态饲用复合酶棕褐色,澄清透明,经过稳定化处理,稳定性高,完全能满足饲料企业运输,贮存和生产需要。本品为生物活性物质,避光、密闭保存,低温4~10℃贮存,保质期半年。
6、颗粒饲料中酶活测定方法
由于饲料成分复杂,磷、糖类背景值高,用常规酶活测定方法难以准确测定,因此制约了酶制剂的推广。曾经一度形成无法比较粉酶、颗粒酶与液态酶效果的局面。康地恩在推行液态酶后喷涂工艺的同时,参考国外文献率先建立了颗粒饲料中植酸酶活性测定方法,在浸提饲料中植酸酶后,用超滤柱滤除游离磷酸根后,按照常规方法测定[13-14]。这对于指导新工艺的改进和普及起到重要作用。
四、饲料酶的体外酶解评价
动物饲养试验周期长,成本高,影响因素多,通过模拟家禽胃肠道环境,建立体外酶解评价体系,可以为饲料酶的使用提供试验依据。
1、木聚糖酶对消化道的适应性
采集国内不同企业生产的木聚糖酶,保存于4℃冰箱。按照KDN企业标准测定酶活(酶活力定义:1g酶粉于40℃,pH=5.0条件下,每分钟水解木聚糖生成相当于1μg木糖还原物质,即为1个酶活力单位,以U/g表示)。
胃蛋白酶耐受性分析:取上述酶的浸体液20ml,调pH 2.6,加入90μl 50mg/ml的胃蛋白酶,在40℃培养箱中温育1.5h,定容至25ml,经过适当稀释后测定残留酶活。
胰蛋白酶耐受性分析:取上述酶的浸体液20ml,调pH 6.5,加入45μl 100mg/ml的胰蛋白酶,在40℃培养箱中温育2h,定容至25ml,经过适当稀释后测定残留酶活。胃-胰蛋白酶耐受性分析:先后进行上述处理后测定酶活。
表1. 模拟肠道环境对木聚糖酶活力的影响(U/g、%)
|
实验组别 |
酶活 |
初始 |
胃蛋白酶组 |
胰蛋白酶组 |
胃+胰蛋白酶组 |
|
KDN |
3496026 |
100 |
30.3 |
38.8 |
36.8 |
|
Y |
1214118 |
100 |
23.8 |
19.2 |
21.1 |
|
X |
3608228 |
100 |
6.2 |
10.9 |
6.9 |
|
J |
3397849 |
100 |
31.0 |
34.4 |
39.2 |
由表1中可知,胃蛋白酶和胰蛋白酶对木聚糖酶的影响相似,且两者先后作用并未进一步影响木聚糖酶的活性。KDN与Y、J对酸、胃-胰蛋白酶耐受性较好,X较差,仅剩余6.9%活性。
2、KDN木聚糖酶对不同饲料原料的体外消化率
模拟体内消化过程,建立四步酶解法,考察KDN木聚糖酶对饲料原料的降解效果,建立小型的酶解数据库。
菜粕、棉粕、花生粕、DDGS、复合底物均研磨,过10目筛。小麦:大豆:玉米= 4:3:3组成复合底物。无酶成品颗粒饲料为肉鸡2号料。对照组(胃蛋白酶﹢胰酶)、实验组(胃蛋白酶﹢胰酶﹢木聚糖酶),二次重复。
1) 准确称取5.00g底物(过10目筛)于150ml三角瓶内,加入30ml去离子水。沸水浴5min,补加10ml去离子水。
2) 冷却后调节pH至2.6,加入胃蛋白酶,使终浓度为1mg/ml,在37℃培养箱中酶解1.5h。
3) 调pH 至 6.5,加入胰酶,使终浓度为2.5mg/ml,转入透析袋内,用15ml pH 6.5的磷酸盐缓冲液冲洗三角瓶,洗液并入透析袋。在37℃培养箱中、盛有4000ml 去离子水的三角瓶中透析2h。
4) 加入木聚糖酶(相当于10万U),37℃培养箱中酶解、透析4h。
测定透析袋内容物及外部透析液中的可溶性戊聚糖(间苯三酚法)和还原糖(DNS法)含量,计算总的释放量。还原糖表征糖的还原末端,包括不同聚合度和不同的糖类,如戊糖、六糖;戊聚糖主要包括不同聚合度的木糖和阿拉伯糖。
表2. KDN木聚糖酶对不同饲料原料体外酶解产生的还原糖和戊聚糖
|
底物 |
戊聚糖(mg/5g底物) |
还原糖(mg/5g底物) | ||
|
对照组 |
实验组 |
对照组 |
实验组 | |
|
大豆 |
38.1 (12.9) |
42.4 (15.5) |
169 (88) |
256 (82) |
|
玉米 |
21.3 (7.8) |
28.1 (7.7) |
1924 (847) |
2484 (876) |
|
小麦 |
49.4 (4.4) |
148.3 (34.4) |
1581 (588) |
1812 (781) |
|
麸皮 |
81.2 (6.1) |
300.6 (81.7) |
434 (185) |
726 (286) |
|
菜粕 |
51 (10.3) |
70.8 (15.8) |
114 (58) |
246 (101) |
|
棉粕 |
60.6 (9.5) |
79 (22.3) |
119 (77) |
168 (101) |
|
花生粕 |
40.6 (15.6) |
51.6 (19.8) |
254 (112) |
372 (144) |
|
DDGS |
50.2 (18.9) |
103.6 (24.7) |
160 (121) |
217 (129) |
|
复合原料 |
47.5 (8.3) |
98 (24.5) |
1248 (524) |
1379 (545) |
|
成品料 |
46.2 (1.9) |
81.6 (22.8) |
409 (197) |
501 (228) |
注:表中括号内数值为外部透析液中的含量,括号前数值为总量。
图1、KDN木聚糖酶对不同饲料原料体外酶解产生的戊聚糖
图2、KDN木聚糖酶对不同饲料原料体外酶解产生的还原糖
图3、KDN木聚糖酶对不同饲料原料体外酶解产生的净还原糖
由图1可以看出,经过消化酶降解,各底物仅产生很少量的小分子戊聚糖透过透析袋,大分子戊聚糖也多在50 mg/5g底物以下。经过木聚糖酶降解后,溶解的大、小分子戊聚糖提高1-3倍,大分子戊聚糖多在50 mg/5g底物以上。尤其含木聚糖较高的小麦和麸皮提高幅度2-3倍,且水溶性小分子戊聚糖提高8倍和13倍,对于麦类降解显著,有助于降低食糜粘度。
图2显示,木聚糖酶作用后,还原糖增加幅度不大,也是麦类提高显著。
如图3,以“试验组小分子还原糖-试验组小分子戊聚糖-对照小分子还原糖”作为水溶性的净的六碳糖,近似认为可被肠道吸收的糖,以提供能量。结果除了小麦酶解产生的净糖增加3%,其余差异不显著。
另外,单一原料的酶解和复合原料的酶解正相关,但没有叠加性,不能依据单一原料的酶解来推测配合饲料的酶解情况。
3、不同木聚糖酶的体外消化比较
考察木聚糖酶(KDN、X和Y)对饲料原料(麸皮、菜粕、棉粕、花生粕、DDGS以及成品颗粒饲料)的降解效果。对照组(胃蛋白酶﹢胰酶)、实验组(胃蛋白酶﹢胰酶﹢木聚糖酶),做一次重复。
准确称取底物5.00g于150ml三角瓶内,加入30ml去离子水。沸水浴5min,补加10ml去离子水,冷却后调节pH至2.6,加入胃蛋白酶,使终浓度为1mg/ml,在37℃培养箱中酶解1.5h。调pH 至 6.5,加入胰酶,使终浓度为2.5mg/ml,转入透析袋内,用15ml pH 6.5的磷酸盐缓冲液冲洗三角瓶,洗液并入透析袋。在37℃培养箱中、盛有4000ml 去离子水的三角瓶中透析2h。加入木聚糖酶浸提液 (相当于10万单位),37℃培养箱中酶解4h。
测定透析袋内容物及外部透析液中的戊聚糖和还原糖含量,计算总量。结果如表4所示。
表3、不同酶制剂对各种饲料原料酶解
|
底物 |
戊聚糖(mg/5g底物) | |||
|
对照 |
KDN |
X |
Y | |
|
麸皮 |
81.2 (6.1) |
300.6 (81.7) |
304.1 (71.5) |
393 (91.9) |
|
菜粕 |
51 (10.3) |
70.8 (15.8) |
53.1 (10.1) |
68.2 (17.2) |
|
棉粕 |
60.6 (9.5) |
79 (22.3) |
62.3 (13.8) |
71.4 (13.9) |
|
花生粕 |
40.6 (15.6) |
51.6 (19.8) |
42.4 (18.7) |
46.6 (19.4) |
|
DDGS |
50.2 (18.9) |
103.6 (24.7) |
59.6 (8.2) |
104.4 (24.3) |
|
成品料 |
46.2 (1.9) |
81.6 (22.8) |
78.5 (14.3) |
94 (18.7) |
|
底物 |
还原糖(mg/5g底物) | |||
|
对照 |
KDN |
X |
Y | |
|
麸皮 |
434 (185) |
726 (286) |
599 (254) |
684 (245) |
|
菜粕 |
114 (58) |
246 (101) |
127 (72) |
160 (89) |
|
棉粕 |
119 (77) |
168 (101) |
121 (75) |
126 (75) |
|
花生粕 |
254 (112) |
372 (144) |
260 (113) |
262 (113) |
|
DDGS |
160 (121) |
217 (129) |
186.5 (108) |
210 (131) |
|
成品料 |
409 (197) |
501 (228) |
426 (184) |
462 (213) |
注:表中括号内数值为外部透析液中的含量,括号前数值为总量。
三种木聚糖酶的体外酶解效果表明,加酶组均显著高于对照,三组很接近,KDN略优。另外,分析戊聚糖这一指标发现,外部透析液中含量比较低,表明试验中所用的木聚糖酶以内切木聚糖酶为主。
结言
酶是饲料消化营养的核心研究内容,植酸酶和木聚糖酶是成功的范例。围绕植酸、糖类、蛋白质的消化酶解,近年来在基因工程、酶工程、发酵工程和体外酶解评价、液体酶稳定性和后喷涂设备等诸多方面都有深入的研究和丰富的文献,新技术的建立和成熟给饲料养殖行业带来福音。液体后喷涂工艺作为颗粒饲料生产工艺的配套技术日趋成熟,尤其适合于禽料生产,本文仅以此为主线作了概要综述,但并非此单一模式。对于饲料原料固有的酶的保持利用、对消化道固有的消化酶的利用、对微生物在液体饲料中的应用等都值得从技术层面研究和经济价值评估。
参考文献
1. Pen J,Verwoerd T C, van Paridon P A, et al. . Phytase-containing transgenic seeds as a novel feed additive for imp roved phosphrous utilization. Bio. Technology, 1993, 11: 811- 814
2. Li J, Hegeman C E, Hanlon R W, et al. . Secretion of active recombinant phytase from soybean cell2suspension cultures. Plant Physiol. , 1997, 114 : 1103 - 1111.
3. 朱连龙, 熊爱生. 植酸酶的生物技术改良及应用. 中国农学通报. 2008, 24:231-236
4. 郦智玉, 孙建义. 基因工程植酸酶表达系统的研究进展. 饲料研究. 2008, 5:19-21
5. 马廷臣. 植酸酶基因工程研究进展. 华中农业大学学报. 2007, 26:730-733
6. 邹立扣,王红宁. 植酸酶及其植物基因工程,微生物学通报. 2005, 32:128-132
7. 杨培龙等: 饲料用非淀粉多糖水解酶转基因植物的研究进展. 中国生物工程杂志. 2005, 25:29-34
8. 姚斌, 范云六. 植酸酶的分子生物学与基因工程,生物工程学报,2000, 16:1-5
9. 张红莲, 姚斌, 范云六. 木聚糖酶的分子生物学及其应用. 生物技术通报. 2002, 3:23-30
10. 龙健儿. β-甘露聚糖酶的研究现状. 微生物学杂志. 1998, 18:44-57
11. 李富伟, 周响艳, 冯定远. 饲用β-甘露聚糖酶的研究及应用. 饲料工业. 2007, 28 :16-20
12. 罗会颖, 姚斌. 来源于Escherichia coli 的高比活植酸酶基因的高效表达. 生物工程学报. 2004, 20:78-83
13. T. W. Kim and X. G. Lei. An improved method for a rapid determination of phytase activity in animal feed. J. Anim. Sci. 2005, 83:1062–1067
14. 刘玉庆, 王海, 孟庆贺. 液态植酸酶后喷涂工艺设备及其产品均匀度测定. 饲料工业, 2007, 28:12-15
上一篇:饲料酶的造粒方法和设备
下一篇:国内外常见酶制剂产品及生产方法(饲料酶)
分享到:
 收藏
收藏
收藏
评论排行
- ·中国发酵企业数据库(4)
- ·(4)
- ·CoQ10高产菌株选育的研究进展(2)
- ·抗生素发酵工艺所用冷却塔的性能分析及处理(1)
- ·微生物菌种选育技术.rar(1)
- ·发酵生产染菌及其防治(1)
- ·赤藓糖醇发酵工艺研究(1)
- ·重组AiiA 蛋白可溶性表达及发酵条件优化(1)
- ·生物反应器设计软件_发酵罐绿色版(1)
- ·酵母粉、酵母浸粉的区别(1)
- ·雷帕霉素研究进展(1)
- ·透明质酸用途和行业概况(1)
- ·黄酒制作工艺(1)
- ·水解(酸化)工艺与厌氧发酵的区别(1)
- ·糖蜜酒精废液处理过程中产生的微生物蛋...(1)
- ·紫杉醇高产菌发酵产物的分离、纯化和鉴定(1)
